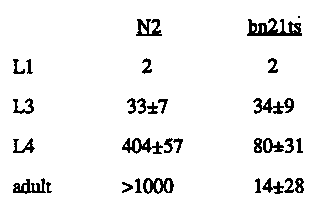Worm Breeder's Gazette 11(5): 87
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
Grandchildless mutations identify loci that encode factors provided maternally and required for normal germ line development. Heterozygous animals produce 1/4 homozygous mutant offspring which are fertile, but which themselves produce all sterile progeny. mes-3 is one of several such maternal-effect sterile loci isolated in the Strome lab (see WBG 11: 3, p. 64). It is defined by three alleles, two non-conditional mutations and one tight temperature-sensitive allele. Four lines of evidence suggest that mes-3 mutations primarily affect germ line development. First, maternal-effect sterile mutations at this locus have been isolated at a rate consistent with isolation of loss-of-function alleles. In addition, mes-3/Df animals display the same mutant phenotype as mes-3 homozygotes, suggesting that the grandchildless phenotype is the loss-of-function phenotype. Thirdly, mes-3 homozygotes grow at the same rate as their wild-type siblings and show little or no embryonic lethality. However, they do have reduced brood sizes, suggesting that their germ lines may be affected, in addition to those of their progeny. Finally, early embryonic lineaging of embryos from mes-3(bn21ts) mothers raised at 25 C indicates that most of the early cleavages occur normally. The only deviation from a wild-type pattern was observed in the P4 division, which took place 5-20 minutes later than in control embryos. The germ line defect of the sterile mes-3 progeny has been analyzed by staining nuclei with DAPI and P-granules with antibody. In wild- type hermaphrodites, L1s hatch with two germ line precursor cells, Z2 and Z3, which proliferate during larval development to a total of approximately 1000 germ nuclei in the adult (not including gametes). Sterile mes-3 hermaphrodite progeny also hatch with two germ line precursor cells, but show a much reduced germ line as adults (see figure). None of the sterile animals produced any gametes and most had normal-looking somatic gonads. In order to determine how animals whose germ lines appear indistinguishable from wild type as L1s develop into sterile adults with reduced or no germ line, an analysis of germ line proliferation in mes-3(bn21ts) mutant progeny was carried out on staged larvae (see table). All experiments were performed at 25 C. In N2 control larvae, Z2 and Z3 began to proliferate late in the L1 stage. By early L3, they contained 33 +/- 7 germ nuclei gonad, increasing to 126 +/- 26 as late L3/early L4s, 404 +/- 57 as late L4s and over 1000 germ nuclei as adults (once again, not counting gametes). In bn21 mutant larvae, Z2 and Z3 also began to divide in late L1, reaching 34 +/- 9 germ nuclei gonad by the mid L3 stage. However, at this point mes-3 proliferation began to lag behind N2 controls. Early L4 animals contained 51 +/- 37 nuclei, and a maximum of 80 +/- 31 was attained in late L4 larvae. After this point, the average number of germ nuclei per gonad dropped, halving by early adult and falling to 14 +/- 28 in older adults. Thus, it appears that loss of mes-3(bn21) function leads to decreased proliferation during later larval stages and loss of germ cells in adults. Initial experiments with the bn35 allele show a similar decrease in proliferation beginning in late L3 animals. However, staged adults have not been tested, so it is unclear whether divisions cease or germ cells are lost in bn35 sterile progeny. Since bn21 is a fully penetrant, fully expressed ts allele ( homozygous mothers produce 2% sterile progeny at 16 C and 100% sterile progeny at 25 C), its temperature-sensitive period (TSP) was determined. Shifting larvae or adults up or down has no effect on the phenotype of the shifted animal, indicating that the TSP occurs during embryogenesis. In order to determine when during embryogenesis, gravid bn21 hermaphrodites raised at 16 C were cut open and staged embryos were shifted to 25 C. When 1-cell embryos were shifted up, all the animals grew up to be sterile, while shifting up gastrulating embryos (2-cell embryos incubated at 16 C for four hours before upshift) resulted in 95% fertile animals. Similar downshift experiments were carried out on embryos. Recovery was never complete, but downshifts of pre-gastrula embryos consistently resulted in 10-15% fertile animals, while downshifts of later-stage embryos resulted in all sterile animals. This indicates that the TSP for the bn21ts mutation is early in embryogenesis, and it is consistent with a role for the mes-3 gene product in early germ line development. Mutations in mes-3 appear to severely reduce the number of germ nuclei present and eliminate gametogenesis in the hermaphrodite. The male germ line does not appear to be as strongly affected (see figure). In male progeny of homozygous mes-3 mothers, the germ line proliferates to a greater extent in at least two of the alleles and some of the males produce sperm. Individual male matings between bn35; him-5 or bn53;him-5 males from homozygous mutant mothers and Dpy odites produced outcross in 10 out of 93 matings. Thus, these males are weakly fertile. Surprisingly, although 7 of the 10 males produced fertile hermaphrodite outcross progeny, three of them produced all sterile hermaphrodite outcross progeny, despite the fact that the mother in all cases was mes-3+. It is worth noting that all three mes-3 alleles are strict maternal effect, and also, that mes-3 homozygous male offspring of heterozygous mothers do not produce sterile progeny when mated with mes-3+ hermaphrodites, so this appears to be a great-grandchildless effect that is passed through the male germ line. In conclusion, the mes-3 locus appears to encode a maternal factor required for normal germ line development. It may function early during embryogenesis, and mutations at this locus cause reduced germ line proliferation, although it does not affect the male germ line as severely as the hermaphrodite germ line. mes-3 maps between unc-38 and dpy-5 on the left arm of chromosome I, so it is on a contig. We have begun molecular analysis of the region and in collaboration with Janet Paulson, are attempting cosmid rescue of bn21ts Hopefully, this will provide insight into the role of the locus in germ line development. [See Figures 1-2]