Worm Breeder's Gazette 11(5): 84
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
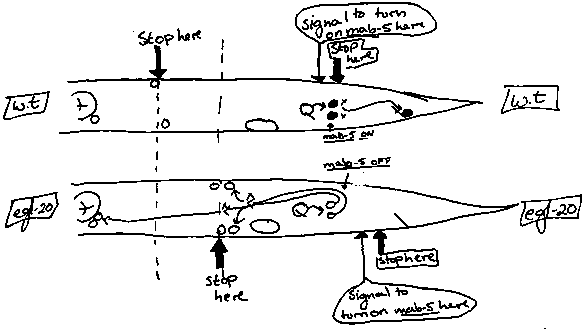
egl-20 initially caught our interest because reduction- or loss-of- function mutations in this gene have two effects on Q cell migration. The most striking Q defect in egl-20 mutants is that the daughters of QL migrate anteriorly, as they do in mab-5(lf) mutants, rather than remaining in the posterior, as they do in wild-type worms. What is unusual about egl-20 mutations is that they also affect the distance the Q descendents migrate anteriorly: in egl-20 mutants the Q descendents Q.paa and Q.pap stop short of their final destination. Genetic and molecular epistasis suggests that the reason the QL.x (i. e. QL.a and QL.p) cells migrate anteriorly in egl-20 mutants instead of remaining in the posterior is because they fail to turn on mab-5. First we found that the mab-5(gf) mutation could suppress the egl-20 Q defect. When the mab-5(e1751gf) mutation is introduced into an egl-20 strain the QL and QR descendents migrate posteriorly. This suggests that egl-20 could act upstream of mab-5 in determining the direction of migration of Q descendants. By examining the pattern of -gal activity in egl-20(n585) mutants carrying an integrated mab-5::lacZ reporter construct (Salser and Kenyon, this WBG) we were able to ask whether egl-20 mutations affect mab-5 gene expression. In a wild-type background QL.x cells stained dark blue 3-4 hrs after hatching in 47/50 worms examined whereas QR.x cells never stained (0/50). In egl-20(n585) mutants carrying the construct, however, QL.x were never observed to stain (0/43). egl-20 mutations do not affect most of the other structures the require mab-5 activity. Consistent with this, the mab-5-lacZ L1 staining pattern did not differ markedly from wildtype, although this has not yet been carefully quantitated. Together these data suggest that egl-20+ is required for mab-5 gene expression in QL.x cells and that QL's descendents migrate anteriorly in egl-20 mutants because they fail to turn on mab-5.The second alteration in egl-20 mutants is a displacement of the Q.paa and Q.pap ( SDQ and AVM/PVM cells). On both sides of egl-20 mutants, these cells are positioned further posteriorly than the normal positions of SDQR and AVM. An additional observation suggests that this alteration is due to an altered signal to stop migrating rather than a simple inability to migrate. In the mab-5(e1751gf);egl-20(n585) double mutant which 'rescues' the QL.x cells and allows them to remain in the posterior, there is a different type of misplacement: this time the Q descendents often migrate too far posteriorly (compared to e1751 alone) . How can we explain the two different effects egl-20 mutations have on the migration of the Q descendents? Possible models which explain the two effects of egl-20:1) 2 separate problems: egl-20 has one activity needed to determine the direction of migration of the Q descendents and another needed for the Q descendents to stop in the right place. It could do this in many ways (interacting with two different proteins, DNA sites, etc., by being present in two forms, one form with two domains, etc....). 2) one underlying problem: a single egl-20-dependent system of positional information is used both to stop migrating cells in the correct place, and also to turn on mab-5. In this model, the wild- type QL cell turns on mab-5 when its initial posterior migration brings it in contact with a cell-extrinsic signal located in the posterior 'mab-5 domain'. In egl-20 mutants this positional information is shifted posteriorly relative to the Q cells, which prevents QL.x cells from receiving their positional cue to turn on mab- 5. Because mab-5 is not turned on, the QL daughters migrate anteriorly. Similarly, when the anterior Q descendents stop short of their full anterior position, it is because the signals to stop migrating are also shifted posteriorly relative to the Q-descendants. In the mab-5(gf);egl-20 double mutant, the extreme posterior displacement of Q.pap and Q.paa can be explained if they too are seeking out their 'proper' final location which the egl-20 mutation has shifted more posterior relative to the rest of the worm. Because mab-5 has been turned on independently of egl-20 by the e1751gf mutation, the cells are able to stay in the posterior region and locate their egl-20-directed final position. A final attractive feature of this model is that it can also accomodate another egl-20 phenotype; namely, altered HSN migration. Like the Q descendants, the HSN cell body is also mispositioned posteriorly in egl-20 mutants ( Trent et al. Genetics 104:619-647,1983; Desai et al. Nature 365:638 646, 1988).