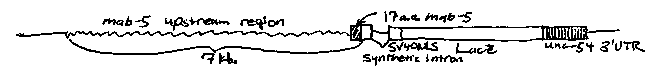
Worm Breeder's Gazette 11(5): 83
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
From the genetic experiments just described we know that mab-5 is necessary and sufficient to make Q.a and Q.p behave in a left-specific manner (remaining in the posterior). Therefore, in wild-type animals the effects of mab-5 must be somehow restricted to act specifically on the QL lineage. Mosaic analysis suggests that mab-5 acts cell autonomously; For the QL migration, mab-5 is required in the descendents of ABp (which include Q and its sister, V5) but not required in V6 or hyp-7 (cells over which the migration takes place). (C. Kenyon, Cell 1986) Assuming mab-5 does act autonomously to control the Q migrations, then some mechanism must restrict mab-5 activity to the QL lineage. Using a mab-5/LacZ fusion we have shown that the mab-5 promoter is specifically active in QL and its descendents. The fusion contains the first 17 amino acids of mab-5 and 7 kb. of upstream sequences fused in frame to Andy Fire's LacZ construct, pD21. [See Figure 1] This fusion was coinjected with the unc-31 marker into unc-31(e169) hermaphrodites, and heritable, unstable lines were obtained. One line became stable after propagation for 6 months at 25 C (which selects for non-Unc-31 animals). The injected markers map to linkage group IV near dpy-20, and have been named muIs2. In animals carrying muIs2 we observe LacZ staining in the QL lineage but never in the QR lineage, consistent with a model in which mab-5 activity is transcriptionally restricted to the QL lineage. We also see strong reproducible staining in the posterior juvenile neurons and posterior intestinal nuclei, whereas less consistent staining is seen in M, V6, and P7/8-11/12. To examine the time course of expression in the QL lineage newly hatched larvae were washed off plates and allowed to develop for various time periods at 25 C before fixation: Faint expression begins about 1 hour prior to the first division just as QL begins to migrate up over V5. LacZ activity increases up to the first division and then remains strong in QL.a and QL.p. After the second division, weak staining is seen in the nuclei of all four cells including the two programmed cell deaths, and by L2 staining in the Q descendents has disappeared. One possibility is that cell- intrinsic mechanisms activate mab-5 expression specifically in QL. The same mechanisms could also cause the initial asymmetry in the migration of QL and QR. Another attractive hypothesis, based on the time of expression, is that the initial migration of QL and QR serves to place them in different environments. The more posterior location of QL could place it in contact with signals that activate mab-5 transcription. A satisfying feature of this model is that the effect of mab-5(lf) mutations on the Q cells can be interpreted as a posterior to anterior transformation rather than a left to right transformation.
