Worm Breeder's Gazette 11(5): 82
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
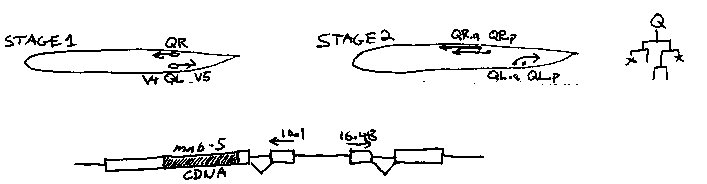
The Q cells are migratory neuroblasts born in symmetric positions on the left and right sides of the animal. Although they divide identically on the two sides, they migrate asymmetrically such that the descendents of QR are found in the anterior, whereas the descendents of QL are found in the posterior. Previous work has shown that there are at least two stages to this migration. Before the first division, QR migrates anteriorly to assume a position just above V4, while QL migrates posteriorly to a position above V5. This phase of the migration is unaffected by mutations in mab-5. Following the first division the cells begin a more complex pattern of migration. QR.a, QR.p, and their descendents continue the anteriorward migration, QL.a and its daughter, QL.ap, continue to migrate posteriorly, and QL.p and its descendents remain stationary. Loss-of-function mutations in mab-5 cause the daughters of QL to behave like the daughters of QR, whereas the putative gain-of- function allele e1751 causes the reverse transformation. [See Figure 1] Expression of a mab-5 cDNA under the worm hsp16.1 heat shock promoter mimics the e1751 phenotype. In these experiments the mab-5 cDNA was inserted into a HpaI site in the second exon of hsp16.1( generously sent to us by Peter Candido.) This construct was injected along with the unc-31 cotransformation marker into mab-5(e2088);unc-31(e169) hermaphrodites, and heritable, unstable lines were obtained. Non-Unc animals were examined starting one hour after hatching and heat shocked on the agar pad (30 minutes at 31 C) just before the first division of Q. In control animals that are unshocked or lack the construct, the descendents of both QL and QR migrate anteriorly as expected in an e2088 background. Eight heat- shocked worms that carry the construct were examined on the left side. QL.a migrated posteriorly over QL.p (7/8 animals), QL.ap migrated posteriorly into the tail (6/8 animals), and QL.p and QL.pa remained stationary (8/8 animals). Although no lineages have yet been obtained for the right side, the QR.paa and QR.ppp cells were found near V4 where QR.p is born (8/8 animals). From this we conclude that mab-5 transcription is necessary and sufficient to make: 1 ) Q.a and Q.ap migrate posteriorly rather than anteriorly. 2) Q.p and Q.pa remain stationary rather than moving anteriorly. We were interested in whether Q.ap migrates posteriorly as a direct effect of mab-5 activity or simply because it is now born close to its new destination in the tail. To address this issue we allowed Q.a to migrate anteriorly past ALM before heat shocking the animals. In 2/2 sides examined Q.ap migrated posteriorly, in one case from V1 back to V3, in another from V3 back to V6. From this we conclude that mab-5 activity is sufficient the make Q.ap migrate posteriorly even if it is born near the head. By carrying out the heat shock at later times in development we have also observed mab-5 activity in the P, V, and M lineages. Heat shock in L3 leads to the production of ectopic rays similar to those seen in e1751.