Worm Breeder's Gazette 11(5): 72
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
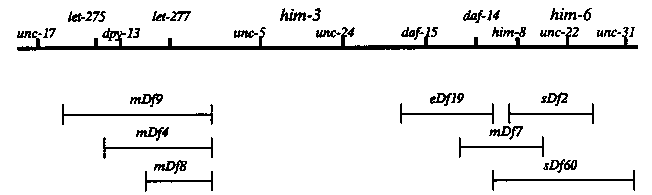
Meiotic mutations in the genes him-3(e1256)IV, him-6(e1423)IV and him-8(e1489)(IV) were originally isolated by Hodgkin et al. (1979) because they caused a high level of X-chromosome nondisjunction. Him- 6 and Him-3 hermaphrodites were found to have elevated levels of autosomal nondisjunction. Him-8 hermaphrodites produced many male progeny due to random disjunction of their X-chromosomes. Unlike the first two mutants, him-8 did not appear to affect autosomal disjunction. I have tested the effects of him-3 and him-6 on recombination frequency in different regions of chromosomes I, III, V and X and tested the effects of all three mutants on the segregation patterns of free duplications. Previous to this study, him-3 and him-6 were known to map on the right end of chromosome IV between dpy-13 and unc-31 (Figure). In order to position them more accurately, him males were crossed to a series of deficiencies from the right end of chromosome IV. him-3 was complemented by all the deficiencies tested. him-6 failed to complement sDf2 and sDf60 but complemented mDf7. This placed him-6 close to unc-22 (Figure). The frequency of males from him-6/sDf60 ( 18/83 = 0.22) and him-6/sDf2 (49/264 = 0.19) hermaphrodites was slightly higher than in him-6/him-6 hermaphrodites (average of 13.7% in recombination experiments). In addition, the number of progeny produced by him-6/sDfs hermaphrodites (279/7 = 40) was less than from him-6/him-6 hermaphrodites (average of 110 in recombination experiments). Because the phenotype of e1423 was more severe when heterozygous to a deficiency, the e1423 mutation may be a hypomorph. Recombination: Compared to the controls, him-3 mutants had a reduced recombination frequency on chromosome I. The reductions were more severe in the bli-3 (25% of control) than in the dpy-5 (68% of control). The severity of the reduction in the bli-3 of him-3 hermaphrodites was confirmed when different markers (let-362 dpy-5) on the left end of chromosome I were used. Similar results were found on chromosomes III where him-3 mutants reduced recombination in the dpy- 17 unc-64 interval (65% of control) and to a lesser degree in the unc- 45 (80% of control). On chromosome V, recombination in the unc-60 was reduced to 50% of the control frequency. On the X-chromosome, him-3 had little effect on recombination frequencies. The recombination frequency in the unc-1 dpy-7 interval was 82% of controls and in the dpy-7 unc-3 interval the recombination frequency 97% of controls. Considering the high level of X-chromosome nondisjunction in Him-3 hermaphrodites, approximately 15% of the progeny from him-3 hermaphrodites were males, the small reduction on X chromosome recombination was surprising. In addition, the lack of an effect of him-3 on X-chromosome recombination differed from the observations made on the autosomes. him-6 also caused reductions in recombination frequency. In the bli- 3 unc-11 region, recombination frequency was reduced in him-6 (75% of control) but not to the degree of reduction in him-3 mutants. There appeared to be no effect of him-6 in the dpy-5 unc-54 region. In contrast, recombination was reduced over the entire lengths of chromosome III and chromosome V. Recombination was also generally reduced on the X-chromosome. Unlike the situation with him-3, the X- chromosome nondisjunction phenotype of him-6 correlated with the reduction in recombination frequency. Autosomal nondisjunction. The level of chromosome I nondisjunction in him-3 and him-6 hermaphrodites was assayed by crossing hT2/+ males to dpy-5; unc-64; unc-36; odites. If a disomy I gamete was produced by the Him strain, then there was a 25% chance it would be fertilized by a I(R)III(R)hT2; IIIN sperm to produce a dpy-5 worm (the I(R)III(R)hT2; III(N) chromosome of hT2(I;III) does not carry dpy-5(+). In the control, hT2/+ males were crossed to dpy-5; odites. No Dpy or Unc progeny were observed in 1882 progeny. Thus, the normal chromosome I nondisjunction frequency was less than 0.002. From the him-3 cross 14 dpy-5 hermaphrodites and 4 Dpy-5 males were recovered in 114 wild-type hermaphrodites and 106 wild-type males. The excess of Dpy-5 hermaphrodites may have resulted from viability differences between hermaphrodites and males or from a segregation pattern such that disomic I gametes were often disomic X as well. The frequency of chromosome I nondisjunction in the Him-3 oocytes was ( 18/238) 0.076. In the him-6 cross, seven Dpy-5 hermaphrodites and five Dpy-5 males were recovered in 471 wild-type hermaphrodites and 451 wild-type males. In this case there was no excess of Dpy-5 hermaphrodites. The frequency of chromosome I nondisjunction in the Him-6 oocytes was (12/934) 0.013. Effects on duplication segregation: We tested the effects of him-3, egregation of chromosomal duplications. In scoring the self-fertilization progeny of hDpx; odites, the him mutants did not increase the frequency of chromosome loss (mitotic and meiotic). The effects of the him mutants on the segregation of duplications from the X chromosome was tested by crossing dpy-5; him-y males to dpy-5; odites. In normal males, hDp12 segregated from the X-chromosome by a non-homologous process (see Herman et al. 1979). him-3 disrupted the nonhomologous segregation of hDp12 resulting in equal numbers of male and hermaphrodite wild-types and Dpy progeny. In contrast, him-6 and him-8 did not effect non- homologous segregation of hDp12 males. hDp31 carries part of the left end of the X-chromosome joined to chromosome I sequences and homologously pairs and recombines with the X chromosome. hDp31 continued to segregate from the X-chromosome in both him-3 and him-8 males, although the efficiency of segregation was reduced compared to controls. The segregation of hDp31 in these strains might be by the nonhomologous process. [See Figure 1]