Worm Breeder's Gazette 11(5): 49
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
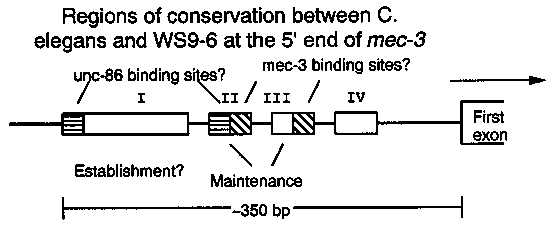
Comparison of the mec-3 DNA sequences from C. elegans and WS9-6 ( another Caenorhabditis species) shows that all the exon sequences are conserved, and in addition, four blocks of nearly identical sequence are found upstream from the start codon (Way, WBG 11, 4). On the assumption that these conserved upstream regions are regulatory sequences, we have used in vitro mutagenesis to investigate their function. Specifically, we have altered various sites in these conserved regions in the mec-3-lacZ fusion plasmid pTU28 (Way and Chalfie, Genes and Dev.3:1823) to see if the conserved sequences are important in determining when and where mec-3 is expressed. Mutations in either Site II or Site III cause a defect in mec-3 maintenance. A disruption of site II causes a severe maintenance defect: the mec-3-lacZ fusion is expressed briefly after the expressing cells are born, then expression is lost for the rest of development. This pattern of expression is the same as when a non- mutant mec-3-lacZ fusion is placed in a strain with a chromosomal mec- 3 mutation. A mutation in site III has a less dramatic effect: expression comes on in all the correct cells, then gradually fades, so that by adulthood, the number of expressing cells is significantly less than for a wild-type mec-3-lacZ fusion. Within the conserved regions are putative binding sites for both mec- 3 and unc-86 (see below and previous newsletters; the mec-3 binding site in Site II is inferred largely from the mutant defect and is only weakly related to the ISL-1 footprint site). Because site II appears to have an unc-86 binding site, we decided to test whether mec-3 expression would depend on unc-86 for maintenance as well as establishment. For ease of strain construction, we first made an integrated derivative of the mec-3-lacZ/unc-22 (antisense) extrachromosomal element uEx4 by gamma-ray treatment (Kari et al. WBG 11, 3, p. 14); this integrated element is termed jeIn2. jeIn2 was placed in a temperature sensitive unc-86 background (n848), so that unc-86 could be inactivated after mec-3 expression had been established. mec-3 does appear to require unc-86 for maintenance. If jeIn2; n848 is shifted to the non-permissive temperature after mec-3-expressing cells are born, they lose expression of mec-3. For example, if this strain is grown at 15 C until the mid-L1 stage, then either stained immediately or shifted to 25 C, the stained L1s express mec-3-lacZ at normal levels. However, if the shifted animals are stained after the L4 stage, expression of mec-3-lacZ is greatly reduced. [See Figure 1]