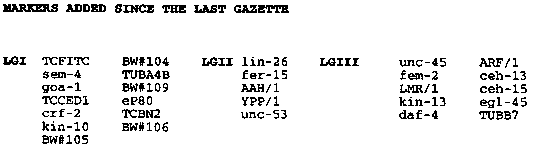
Worm Breeder's Gazette 11(5): 15
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
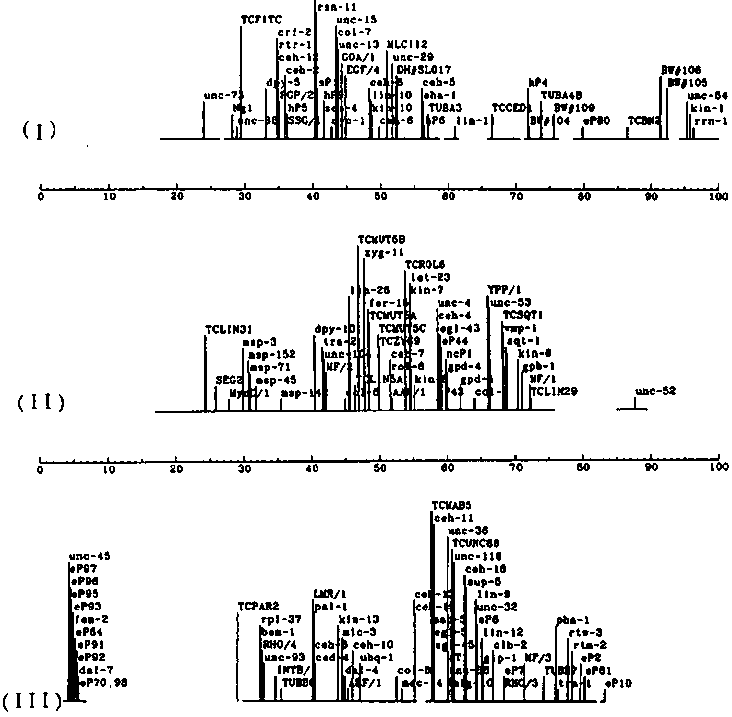
After the introduction of YAC libraries into the C. elegans mapping project, we pursued reciprocal hybridization of YACs and cosmids to link together the cosmid contigs (see Coulson et al., Nature 335, 184- 186). The YAC libraries have been increased to over 9000 clones, with over 3000 used in hybridization. A steady reduction in contig number resulted so that by the beginning of this year the number of contigs had fallen to 170 (from a starting number of 700) and the average size had risen from 125kb to more than 550kb. This greater continuity, combined with the ever increasing number of genetic loci placed on the physical map, increased the amount of DNA positioned on the genome map from less than 20 Mb to more than 75 Mb. The YAC coverage of the contigs was almost complete, and less than 1000 were required to give redundant (2 fold) coverage of the physical map. A grid of these clones (genomically ordered at the time of selection) has been prepared on a postcard sized piece of nylon (958 clones in all) and replicas made for distribution. Using the key provided and the electronic map (see WBG#10;3) it is now a straightforward procedure for a lab to locate any given DNA sequence on the genome map. They have been used successfully in a number of labs. These 'polytene' filters may be obtained from either St. Louis or Cambridge by request. By the beginning of the year, it had become clear that the reciprocal hybridization strategy was decreasingly useful and another approach was needed. The remaining contigs were largely of two classes: (1) contigs with YACs as the likely endmost clones; and (2) small contigs lacking YACs. The contigs with YAC ends were blocked from growth because the reciprocal hybridization strategy provides no way to recognize YAC-YAC overlaps. To obtain an end probe from the YACs, sequence was obtained via an adaptation of Engelke's genomic sequencing method (Methods in Enzymology). Primers were made and the PCR product used to probe to the YAC grids. Hybridizing YACs were tested with the same primer pairs by PCR to confirm the overlap. A total of 34 joins and rearrangements have resulted from the analysis of 186 ends. Because these joins involve just the ends of YACs they are particularly vulnerable to artifact; please inquire if you need to know the status of particular areas. The small YACless contigs were of a variety of types. Some contained only cosmids made in the pJB8 vector that had been left out of the above grids because sequences share with the YAC vector. Some had repeated sequences that led to ambiguous positioning. Others simply had failed to hybridize with YACs tested. To assign these cosmids to YACs, end sequence was obtained by direct sequencing of cosmid DNA, PCR was used to generate small probes and positive YACs were tested by PCR to confirm assignment. Of 50 cosmids from which we have useful data, 38 have been assigned to YACs. Currently, for 31 cases these YACs either had previously been placed on the physical map or have been subsequently linked to larger contigs, thereby locating the small contigs. Many of the remaining clones failed to give a PCR- product from nematode DNA and have been set aside. Only 2 clones were found which gave a PCR product from worm DNA and failed to identify any overlapping YAC. Together these efforts have reduced the contig number to 102. With the joins obtained and the continuing efforts of the community to clone and assign genetically mapped fragments on the physical map, 85 Mb of DNA is now aligned with the genetic map. [See Figures 1-4]