Worm Breeder's Gazette 11(4): 98
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
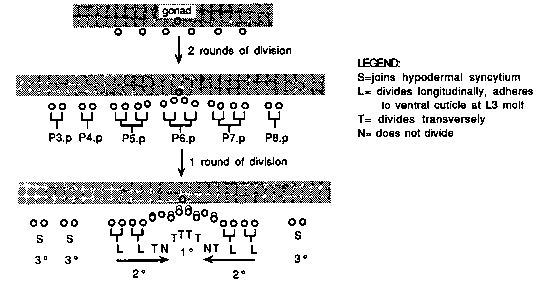
The vulva of C. elegans is formed from the descendants of six vulval precursor cells (VPCs), P(3-8).p. The 2 VPCs, P5.p and P7.p, produce asymmetric lineages of fixed polarity. We are investigating the mechanisms by which the polarity of the 2 lineages is established and oriented. The best characterized indicator of 2 lineage polarity is the orientation of the final nuclear divisions and subsequent migrations, which produce characteristic arrangements of the daughter nuclei. [See Figure 1] In adults, the pattern of lectin staining indicates 2 polarity ( Link, et. al., Development 103: 485-495, 1988). Normally the cells proximal to the P6.p descendants stain with wheat germ agglutinin (WGA) , while the distal (L) cells do not; and, as discussed below, the pseudovulvae of lin-12(d) mutants stain with characteristic patterns ( G. Freyd and R. Horvitz, pers. comm.). Genetic analysis. Mutations in the lin-18 gene abolish or reverse the polarity of the 2 lineages. The phenotypic consequence is the presence of two ventral protrusions (bivulva), or a single large protrusion at the vulva. The two existing alleles of lin-18 are incompletely penetrant; some animals have normal 2 lineages. At the plate level, about 33% of lin-18(e620) hermaphrodites are bivulva. Most of the remaining animals have a protruding vulva, and often rupture at the vulva, suggesting that the e620 defect affects other 2 - specific functions in addition to perturbing the axes of nuclear division. We are studying lin-18 to learn its role in specifying polarity. Since the existing lin-18 alleles are incompletely penetrant, we have begun by screening for deletions of the lin-18 locus, to help define the null phenotype. We have isolated a deficiency, syDf1, which covers at least the region from unc-78 to dpy- 8, including the lin-18 locus. syDf1/lin-18(e620) animals are viable. Experiments are in progress to determine whether the deficiency increases the penetrance of the e620 phenotype. We have also constructed congenic strains as a first step to cloning this locus by correlating the genetic and physical maps. Ablation experiments. We wish to learn how signals from nearby cells might influence the polarity of the 2 lineages. To do so, we are using lin-12(n137) animals, in which all six VPCs form 2 lineages, whether or not inductive signal from the gonad is present. This fact allows us to perform cell ablation experiments to examine the role of the gonad, and of neighboring VPCs, in specifying 2 lineage polarity. By Nomarski observation of n137 animals, P(3-5).p lineages typically are oriented to the posterior, P6.p to the anterior, and the polarities of P7.p and P8.p are variable. n137 adults typically have five to six ventral protrusions (pseudovulvae). About 80% of the time, the P5.p and P6.p descendants appear cooperatively to form one pseudovulva larger than those formed by the single lineages. WGA usually stains half of a pseudovulva formed by a single lineage, and the central part of the large pseudovulva formed by P5.p + P6.p lineages. We score this as posterior staining and anterior staining for the P5.p and P6.p lineages, respectively. Preliminary observations indicate that WGA staining frequently but not invariably correlates with lineage polarity by Nomarski. We are investigating this correlation more thoroughly. WGA staining patterns for nonablated and ablated n137 animals are shown in the following table ( 'apolar' = stained at the tip, or stained all over; 'inconclusive' = pseudovulva did not evert) : [See Figure 2] All lineages were oriented to the posterior in all of 5 gonad- ablated worms lineaged by Nomarski. These observations indicate that the gonad does influence the polarity of the P(6-8).p lineages. For example, WGA staining of the P7.p pseudovulva in nonablated animals is mainly anterior, while in gonad ablated animals it is mainly posterior. This is somewhat surprising, since n137 animals lack an anchor cell, which is required for vulval induction in wild type animals. It possible that there is still a low level of anchor cell signal produced, even though the anchor cell itself does not differentiate. Alternatively, the 2 lineages may orient to a signal from other cells in the gonad. The products of the Vul genes lin-2, lin-7, and lin-10 are probably not necessary for the effect, since the polarity of P6.p and P7.p is variable in n137; Vul double mutants (Sternberg and Horvitz, Cell 58: 679-693, 1989). We are performing further ablations to explore these possibilities, and to examine whether VPC-VPC interactions play a role in orienting lineage polarity.
