Worm Breeder's Gazette 11(4): 93
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
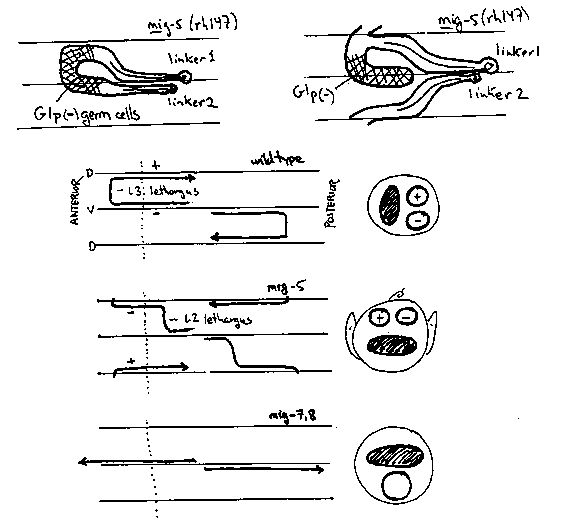
GONADOGENESIS mig-5 is required for determination of distal tip cells (DTCs) in both sexes and for their program of migration in hermaphrodites. In a sample of rh94 hermaphrodites (N=119), about 10% (23/238) of the DTCs were absent, including 1 sterile individual with no DTC and 2 individuals with three DTCs. In rh147 (N=101), about 24% (49/202) of the DTCs were absent, including 6 individuals lacking both DTCs. These frequencies of double DTC loss, 1% = 10% X 10% and 6% = 24% X 24%, are consistent with independent behavior of gonadal precursors Z1 and Z4. Using the more penetrant allele rh147, we hope to follow an aberrant gonadal lineage through the early larval stages. A simple possibility is that presumptive DTCs sometimes become sheath/spermathecal precursors like their sister cells; rare extra DTCs in rh94 could reflect a converse transformation. Conceivably, null mutants of this gene may make no DTCs. Although some rh94 and rh147 males have normal gonads; most lack a functional DTC and are hence defective in germline proliferation (Glp). Such sterility is much commoner in mig-5 males than hermaphrodites perhaps because DTCs arise by a somewhat different lineage in males (Z1.a/Z4.p) than hermaphrodites (Z1.aa/Z4.pp)(1). Sterile mig-5 males often have a second linker cell. In wild type, both Z1.paa and Z4.aaa are competent to become linker cells (primary fate) but cell interactions divert one cell to a secondary fate (vas deferens precursor). By analogy to the hermaphrodite anchor cell, this interaction may require direct cell-cell contact (2). In mig-5, the pattern of cell contacts may be perturbed, allowing both cells to become linker cells. Invariably, one linker cell has a normal trajectory but the second cell may migrate less completely (Figure 1). In hermaphrodites, distal tip cells (DTCs) migrate from ventral to dorsal body muscles during L3 lethargus. In some mutants, dorsal movement occurs precociously (dpy-24) or fails entirely (mig-7, rors give a distinctive gonadal shape that is recognized at low magnification with bright field optics. In rh94, 5- 10% of DTCs execute their dorsal movement during L2 lethargus, i.e., a full larval stage early (Figure 2). Q MIGRATIONS Eight known genes (emb (e1933; III), hch-1(X), mab-5(III), mar(rh148; X), mig-1(I), mig-5(II), mup(rh80;X),and unc-40(I)) are required for the posterior migration of neuroblasts QLa and QLp during the first larval stage (2,3; unpublished data). Each of these mutants has unique, additional phenotypes, e.g., hch-1 affects secretion of a protease used for digesting the eggshell. emb(e1933) and mig-5 have strong maternal effects: emb+ expression is sufficient for embryogenesis but not larval Q migrations while maternal mig-5+ expression is sufficient for larval cell lineages including Q migrations. For most of these mutants, including mig-5 (rh94), the QLa/QLp defect but not other phenes, is suppressed in doubles with the semidominant mutation e1751 causing posterior migration of neuroblasts QRa/QRp when alone. The major changes in cell direction during gonadal cell migrations may be caused by intrinsic changes within the migrating cells (E. Hedgecock, D. Hall, J. Culotti, in preparation). In this view, the evolving trajectory reflects progressive differentiation of the motile cells. Mutations affecting the Q lineage/migration suggest a slightly different emphasis; here, changes in cell direction coincide with cell divisions and determination of new cell types. By analogy, gonadal migrations may be regarded as degenerate cell lineages with periodic changes in cell direction (alias type) but no actual divisions. mig-5 affects various cell fates in the larval gonad and hypodermis; like mab-5, it may be affecting cell migrations indirectly by regulating genes involved in cell adhesion and guidance (4,5). [See Figure 1]