Worm Breeder's Gazette 11(4): 87
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
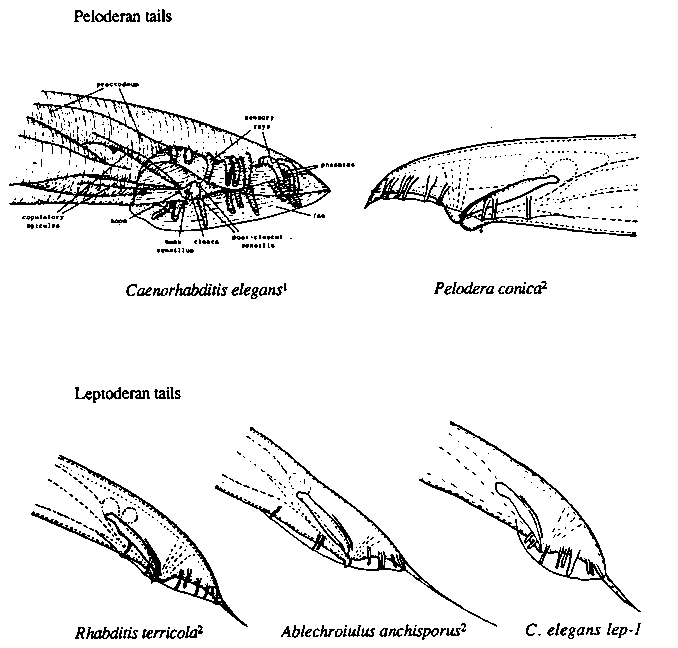
Morphogenesis plays an important role in the formation of the C. elegans male tail. Just before the L4 larval molt, morphogenesis of the male bursa results from retraction of the most posterior cells. In the wake of the retraction, the outer cuticle folds and flattens into a fan-like structure, the bursa. As another part of this retraction, the syncytium, hyp10, forming the tail spike of the earlier larval forms, shrinks and does not protrude posterior of the bursa (i.e., forms a peloderan tail--see Figure). Three mutations isolated in our laboratory, bx37, bx42, and bx73, cause failure of only the hyp10 syncytium to retract in males, leaving a tail spike (i.e., a leptoderan tail--see Figure). No other defects appear to be associated with this phenotype; males are fertile and mate efficiently. Alleles bx37 and bx42 are semidominant ( semidominance has not yet been tested for bx73). As homozygotes, bx37 and bx42 are 100 and 92% penetrant, respectively; as heterozygotes, about 5% of the males possess leptoderan tails. These alleles both show variable expressivity; tail spike phenotypes range from a schlerotic bump to a spike as long as the bursa itself. bx73 males show the highest expressivity. Alleles bx37 and bx42 fail to complement and map between positions +7.4 (2-factor data) and +11.4 ( left of eDf3) on LGI. No mapping or complementation data have yet been obtained for bx73. Based on the frequency of their recovery, we suggest that these mutations are nulls or hypomorphs, semidominance being explained by haploinsufficiency. bx37 and bx42 therefore define a gene (designated lep-1, for LEPtoderan) required for hyp10 retraction during male tail morphogenesis. In one model, reduction or loss of lep-1 activity would result in sexual transformation of hyp10 toward the hermaphroditic fate. It is noteworthy that male members of several genera confamilial with Caenorhabditis (e.g., Rhabditis) possess leptoderan tails, implying that the gain (and/or possibly the loss) of male tail spikes has probably occurred several times during the evolution of rhabditid nematodes. It is likely that tail spikes in these species arise not through cell lineages peculiar to certain leptoderan species, but from the failure of the most posterior cells to retract, as in lep-1 mutants of C. elegans. Similarly, the Rn cell lineages in the male tails of Panagrellus redivivus and C. elegans are identical, yet fan morphogenesis does not occur in P. redivivus. Although completely fanless mutants have not yet been obtained in C. elegans, mab-17( e2167) males fail to stabilize the retraction of hyp7 and other cells normally participating in this process. We suggest that lep-1 and mab- 17 are members of a class of 'heteromorphic' genes which have been evolutionarily involved in producing different forms, not by changes in cell lineage patterns (as, for example, with heterochronic genes), but by changes in morphogenetic processes. Of interest to taxonomists is the fact that absence of a tail spike is the primary distinguishing character of the rhabditid subfamily Peloderinae, to which Caenorhabditis belongs. In light of the present data, that this structure can be gained (and possibly lost) by a mutation in a single gene, a reconsideration of the use of some morphological features in rhabditid taxonomy may be warranted. [See Figure 1]