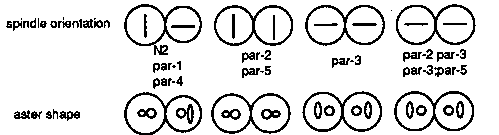
Worm Breeder's Gazette 11(4): 81
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
In an effort to understand the relationships among the par genes we have been analyzing double mutant combinations. The phenotypes of mutants in the five par genes are similar but readily distinguishable from one another. Strong loss-of-function mutations in all genes affect cleavage spindle position and orientation, cleavage rate, P granule localization, and development of the intestine but do so in a gene-specific manner. We have interpreted these phenotypes to indicate that the par genes encode components of a maternally provided system for establishing cleavage asymmetry and cytoplasmic localization in the early embryo. The results from the five doubles we have examined thus far indicate that the genes are not in a simple dependent pathway but they do interact. There are no clear epistatic relationships, although individual aspects of the phenotypes may show epistasis (see below); the cellular phenotypes of the strong mutants are simply additive, not synergistic. However, we often see synergism with respect to viability in double mutant combinations of weak alleles. The most striking example is the double homozygote par-2(it5ts); 7ts). it5 gives greater than 70% viable progeny at 15 C and it57 gives nearly 100% viable progeny at 15 C. it5;it57 doubles give almost complete lethality at 15 C. We also see dominant enhancement of weak alleles of one par by one copy of a strong allele at a second par gene. Two particularly interesting double mutant combinations are par-2 par-3 and par-3;par-5. par-3 affects the orientation of the second cleavage spindles in a way that is opposite to par-2 and par-5.[See Figure 1] We were surprised to find that par-3 is epistatic to par-2 and par-5 for this particular phenotype. This indicates that none of the three genes are required for the rotation of the P1 spindle. Rather, par-3+ acts to prevent spindle rotation and par-2+(5+) acts to insure that par-3+ acts only in the AB. This could result either from localization of par-3+ activity to the AB or by action of par-2+(5+) in the P1. We think it likely that par-2+(5+) acts in the P1 to inhibit or modify the action of par-3+, because we have obtained evidence that par-3+ activity is present in P1. In the wild type, P1 spindle rotation occurs reproducibly in embryos that are slightly flattened ( squashed) by pressure from the coverslip. Our initial observations of par-3 mutants were done on unsquashed embryos. When we observe par-3 mutant embryos under squashed conditions, the orientations of AB and P1 spindles are much more variable than under unsquashed conditions. We conclude, therefore, that although the rotation of the P1 spindle does not absolutely require par-3+ the strength of the rotation or the stability of the new spindle position is dependent upon par-3+ function. Perhaps par-3+ encodes an element of the cytoskeleton that normally stabilizes spindle position in the early embryo. par-2+ then acts in the P1 to modify the par-3+ activity allowing the spindle to rotate. Another aspect of the par phenotype may relate to this point. In the wild type, the asters of the first cleavage spindle show different behavior in P1 and AB. In the P1 cell the aster becomes disc shaped while in the AB the aster remains spherical. In par-1 and par-4, the P1 aster becomes disc shaped. In par-3, both AB and P1 asters become disc shaped, although not to the same extent as in wild type. In par-2 and par-5 both asters remain spherical. In the par-2 he asters are disc shaped (par-3;par-5 has not yet been checked). Thus the change in aster shape at the end of the first division seems to correlate with spindle rotation.