Worm Breeder's Gazette 11(4): 70
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
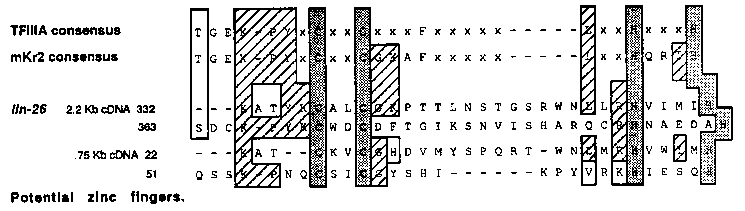
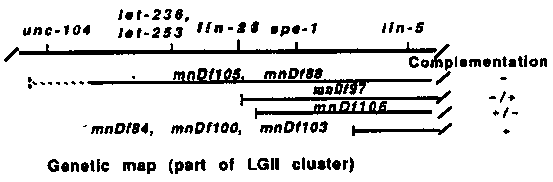
During development, many cells divide asymmetrically to produce daughters that differ in their fates. The mutation lin-26(n156) II disrupts the asymmetry of some cell divisions. This mutation is the only lin-26 allele: it is recessive and is probably not a null allele since it results in lethality in trans to a deficiency (1). lin-26 hermaphrodites are vulvaless. More specifically, the 12 Pn.p cells. which include the six P(3-8).p vulval precursor cells, appear to be transformed to express the fates of their sisters, the 12 Pn.a neuroblasts (2). In wild-type animals, the Pn.a cells undergo up to three rounds of divisions and generate five neural descendants, one of which, the VD neuron, expresses the neurotransmitter GABA. In lin-26 animals, the Pn.a cells are unaffected while the Pn.p cells undergo up to three rounds of divisions and generate descendants with a neural appearance, including extra GABAergic cells (3; Steve McIntire personal communication). These data suggest that lin-26 functions in making the Pn.p cell fate different from the Pn.a cell fate. We have started a molecular analysis of lin-26. The mutation n156 was mapped relative to a set of seven deficiencies spanning part of LGII. The deficiencies mnDf84, nt n156, the deficiencies mnDf97 and mnDf106 partially complement n156, and the deficiencies mnDf88 and mnDf105 do not complement n156 (see genetic map). We physically mapped the left breakpoint of these seven deficiencies using cosmids from the contig spanning most of LGII. The overlapping cosmids C24G10 and C09G7 identify the breakpoint of the deficiencies mnDf97 and mnDf106 (either as polymorphisms or as missing fragments). Using germline transformation, we have been able to rescue n156 by injecting the cosmid C18C9. which overlaps with C24G10 over a 10 Kb region. Subsequent subcloning identified a 9 Kb region sufficient to almost fully rescue n156 (the vulva is slightly protruding, whereas if this 9 Kb fragment is extended on one side the animals are phenotypically wild-type). Northern blot analysis shows that three overlapping RNAs, of 2.2 Kb, 1.5 Kb and 1.45 Kb. are transcribed from the central part of this 9 Kb region. The 1.5 Kb and 1.45 Kb RNAs hybridize to the same set of sub-fragments from the 9 Kb region; most likely they differ by the presence of a small exon or the choice of different 5'/3' ends. We have isolated 14 cDNAs (from 500,000 plaques screened using Stuart Kim's cDNA library): one is a 2.2 Kb cDNA and corresponds to the 2.2 Kb transcript, and one is a 0.75 Kb cDNA and corresponds to the 1.5 Kb/1.45 Kb transcript. The 2.2 Kb cDNA can potentially encode a protein of 473 amino acids. The incomplete 0.75 Kb cDNA contains an open reading frame (ORF) of 131 codons; the 3' untranslated region of the latter cDNA overlaps with the 5' untranslated region and beginning of ORF of the 2.2 Kb cDNA. Database searches (TRANSGEN) indicate that both cDNAs could encode a protein displaying two potential zinc fingers related to, although different from, the Kruppel/mKr2 type of zinc fingers (4; see Figure). We do not yet know which of these three RNAs (if any) are required for lin-26 function. The likely existence of three lin-26 transcripts suggests that alternative splicing and/or alternate use of promoters/terminators plays a role in the expression of lin-26.[See Figures 1 & 2]