Worm Breeder's Gazette 11(4): 110
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
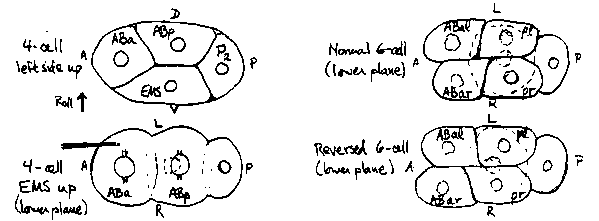
Both the embryo and the embryonic cell lineage are asymmetric from the 6-cell stage onward, particularly with regard to the ABa lineages ( 1). The general bilateral symmetry of the animal is superimposed on the asymmetric lineage by lineally nonhomologous cells adopting analogous fates at equivalent contra lateral positions during embryogenesis (1), but several asymmetries remain throughout larval development, such as the anterior-posterior positions of the left and right coelomocyte pairs (2) and the anterior-r, posterior-l orientation of the primordial and developing gonad (3). These asymmetries normally have the same handedness in all individuals; in N2 populations grown at 16 C, no animals with reversed gonad handedness were found among about 2500 examined. Handed asymmetry first becomes evident in the embryo between the 4- and 6-cell stages, when skewing of the l-r cleavages of ABa and ABp results in positioning of the al and pl daughters anterior to ar and pr, respectively, with al more ventral than ar [see Wood, WBG 11,#3(May) :73, 1990]. Reversal of handedness was accomplished using methods similar to those of Priess and Thomson (4), by rolling a 4-cell embryo to ventral side (EMS) up with a microneedle and then pressing down and back on the left ventral surface of ABa during AB-cell cleavage. When successful, this procedure reverses the normal skewing of both AB spindles and results in ar and pr being anterior to al and pl, respectively, at completion of the cleavage (see diagram). The embryonic development of a reversed embryo was observed and recorded using the '4D Microscope' developed by J. White. (This consists of a Nomarski microscope equipped with a video camera connected to a computer controlled optical disc recorder and focussing drive motor, which automatically records complete sets of serial optical sections at predetermined intervals and allows subsequent playback of consecutive images from any desired optical section to facilitate lineaging.) Cell assignments in the reversed embryo, based on timing of divisions, relative positions, and lineaging to confirm identities of AB descendants, were entirely consistent with a completely enantiomorphic but otherwise normal cleavage pattern. For example, MS descendants were located on the left side of the early embryo and C descendants on the right; MSa and Ca descendants were to the right of MSp and Cp descendants, respectively, and so on. In the L1 that hatched from this embryo, 14 normally asymmetrically placed nuclei scored were all l-r reversed: Z1, Z2, Z3, and Z4 in the gonad primordium, M, the four cc's, Q2, the exc cell, mu sph, hyp11, and PVR. The adults that developed from other reversed embryos showed reversed handedness for the somatic gonad, placement of coelomocytes, and the positions of neuronal cell bodies relative to processes in the ventral nerve cord. The reversed animals were healthy, moved normally, and were normally fertile. These results indicate that ABal is equivalent to ABar and ABpl to ABpr, and, therefore, that the left-right differences in lineage patterns and cell fates exhibited by descendants of ABa and ABp must be dictated by cell interactions, which differ on the two sides as a consequence of the asymmetric positioning of these cells relative to others in the embryo. The respective cell contacts of the ABal-pl and ABar-pr pairs are still equivalent in the 8-cell embryo despite the asymmetry, and earlier laser ablation experiments (1) provide evidence for cell autonomy of many AB-cell fates subsequent to the 51-cell stage. Therefore, the determinative interactions are most likely to occur between the 15-cell and 51-cell stages. [See Figure 1] Thanks to A. Chisolm, J. Rothman, J. Sulston, and J. White for timely coaching and sage advice.