Worm Breeder's Gazette 11(4): 103
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
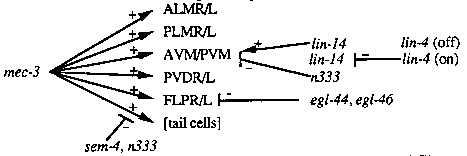
mec-7 is a -tubulin gene that is specifically expressed in the six touch receptor neurons (ALMR/L, PLMR/L, AVM, PVM; WBG 11#3, 38-39). We have used mec-7 in situ hybridization to look for changes in mec-7 expression in over 50 mutant strains that are known to have altered neuronal lineages. We previously reported changes in mec-7 expression in lin-32, mec-3, lin-14, These data and the results reported here support a combinatorial model in which the action of several genes, none of which are expressed in a cell specific fashion, results in the cell-specific expression of touch cell features. Both positive and negative regulation appears to be present. Three (or possibly four) additional genes change the pattern of mec- 7 expression. In addition to the six touch cells, two additional cells are mec-7-positive in egl-44(n1080) and egl-46(n1075, n1076, n1127) mutants in more than half animals examined. These extra cells are found near to where the FLP cells normally differentiate, and we have provisionally identified them as such. The FLP cells (and PVD cells) are neurons that also express a mec-3-lacZ fusion (Way and Chalfie, 1989, Genes & Dev. 3, 1823-1833). sem-4(n1971) animals have an extra pair of mec-7-positive cells in the tail in all animals examined. These cells lie close to the PLM cells, but we do not know which cells they are. They could be the same cells reported by Jeff Way to have -galactosidase activity in a partially deleted mec-3-lacZ fusion (WBG 11#2, 24). Nonetheless, these data suggest that the wild-type sem-4 gene prevents mec-7 expression in these cells. An attractive hypothesis is that sem-4 may negatively regulate mec-3.lin-29(n333) animals also occasionally have two extra mec-7-positive cells in the tail as well as an extra set of AVM/PVM-like cells. Since other lin-29 alleles (n836, n1440) do not have this phenotype we do not know whether n333 is an unusual lin-29 allele or whether the lin-29 strain harbors a second mutation. We have crossed n333 animals with wild-type males and are testing the subsequent outcrossed progeny using in situ hybridization. At this time we do not know how direct or indirect action of these genes is on touch cell differentiation. Nonetheless our results can be diagrammed as follows to show how the combinatorial action of these genes might control cell fate: [See Figure 1] Although lin-14 is expressed in the ALM cells (Ruvkun and Giusto, 1989, Nature 338, 313-319), mutants with lin-14 loss-of-function alleles are not touch insensitive. Our results would suggest that an additional factor, perhaps acting with the lin-14 product, is compensating the phenotype in lin-14 loss of function mutants and results in normal ALM and PLM differentiation into touch cells.