Worm Breeder's Gazette 11(4): 101
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
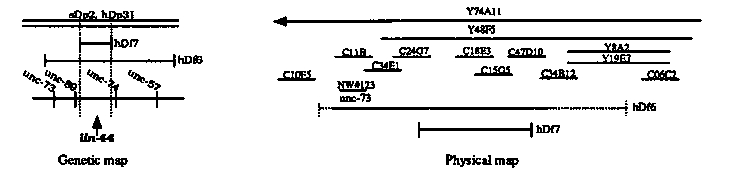
During C. elegans development the two sister cells generated by most cell divisions differ in their fates. Mutations that cause such normally asymmetric cell divisions either to become symmetric or to reverse the polarity of their asymmetry may identify genes responsible for the control of asymmetric cell divisions lin-44.Last year we reported the isolation of a mutation defining a new gene that affects certain asymmetric cell divisions (Herman and Horvitz, CSH C. elegans Meeting Abstracts 1989, p. 277.) The mutation lin-44(n1792) primarily causes certain asymmetric cell divisions to be reversed in polarity. The B, U, F and T cell lineages are all affected in males, resulting in defective male tails. The T lineage is also affected in hermaphrodites, causing the phasmids (sensory structures in the tail) in both sexes to fail to fill with fluorescent dyes. In addition, some hermaphrodites are egg-laying defective. Using a non-complementation screen, we have obtained another lin-44 allele, n2111. The n2111 allele behaves similarly to n1792 when homozygous and when heterozygous with the wild-type allele or with a deficiency. Also, both homozygotes look similar to n1792/Df and n2111/Df heterozygotes and to the n1792/n2111 heterozygote. Thus both n1792 and n2111 appear to be reduction-of-function alleles, and may be null alleles. These two mutations are different since n1792 is suppressed by one copy of the amber suppressor sup-7(sb), while n2111 is not suppressed. The cells affected by lin-44 are related by their position in the tail and not by lineage, which suggests that cell-cell interactions play a role in controlling these asymmetric cell divisions. In his studies of mab-9, Andrew Chisholm observed that when the B cell is killed in wild-type males, the polarities of the F and U cell lineages are reversed, just as they are in lin-44 animals. This observation suggests that lin-44 may function in cell-cell signalling between B and F and U. We are trying to determine which cells, if any, can influence the polarities of the B and T cells. We have killed the B cell in wild-type L1 males and looked for effects on the polarity of the T cell lineage and have killed T in wild-type L1 males and looked for effects on B. We have not seen an effect in either case. Furthermore we have killed the cell AB.prpaaaap, the mother of the B cell, in him-5 and lin-44; ryos and have not observed an effect in either case. Since the B cell never existed in these animals, this result strongly indicates that B does not influence the polarity of T. We are performing similar experiments by killing the T cells in the embryo before the B cell is born. We have not observed an effect on the B cell in him-5 males. Studies of lin- 44; e in progress. Finally, we are cloning the lin-44 locus. lin-44 maps about 0.1 m.u. to the right of unc-73 I, which has been cloned in the laboratory of Joe Culotti . The unc-73 clone identified a 1,100 kb contig. We mapped two deficiencies that uncover lin-44, hDf6 and hDf7, to the unc- 73 contig by probing Southern blots of N2, hDf6 dpy-5 unc-13; hDp31, and hDf7 dpy-5 unc-13; sDp2 DNA with labeled cosmids on the contig. By comparing band intensities between wild-type and deficiency-bearing strains, we determined whether a particular cosmid was under the deficiency. We also looked for polymorphic bands in the deficiency strains, indicating a possible breakpoint. From these experiments we have generated the physical map shown below. We plan to isolate a clone containing the lin-44 gene by microinjection of cosmid or YAC DNA. [See Figure 1]