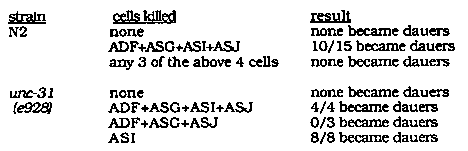
Worm Breeder's Gazette 11(1): 52
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
Dauer larva formation is regulated by the presence of food and dauer pheromone in the worm's environment. Wild-type C. elegans animals become dauer larvae inappropriately (in the presence of bacteria) if four sensory cell types, the amphid cells ADF, ASG, ASI, and ASJ, are killed with a laser microbeam (WBG 10: 2, p.42). We infer that these sensory cells recognize either food, pheromone, or both stimuli. Leon Avery has described a mutation, aex-3(ad418) X, that has a synthetic dauer constitutive phenotype in combination with unc-311e928) IV ,WBG 10: 2, p.39). aex-3 also has a pharyngeal pumping defect in combination with unc-31; de redundant functions in these two processes. We thought the redundancy between these two genetic functions might correspond to the redundancy among sensory neurons controlling dauer formation. To test this idea, we killed sensory neurons in unc-31 animals and observed the development of the altered animals in the presence of plentiful bacteria. Selected data are shown below: [See Figure 1] Only one of the four cells that can prevent dauer formation in N2 seems to be active in unc-31, since the death of ASI is sufficient to lead to dauer formation. We conclude that ADF, ASG, and ASJ are functionally inactivated in unc-31 animals. This could be due to a requirement for the unc-31 gene product either in the sensory cells or in cells that act as their targets in dauer formation. The suggestion that unc-31 is required for sensory neuron function is supported by our observation that unc-31 animals are unable to chemotax to aqueous (Cl-, cAMP) or volatile (benzaldehyde, isoamyl alcohol) attractants. Since killing ADF, ASG, and ASJ does not lead to chemotaxis defects of that severity, it seems likely that additional cells are affected by unc-31 mutations. Alberts and Riddle (J. Comp. Neurol. 219:461 (1983)) have described a remodeling of the amphid sensilla in dauer larvae. As a result of this remodeling, the sensory neurons ASG and ASI are not exposed to the environment in dauer larvae, although they are in normal larvae and adults. This could render those neurons insensitive to external stimuli in dauer larvae and therefore unable to mediate recovery from the dauer stage. Indeed, we have found through laser experiments that only ASJ and possibly ADF appear to affect recovery of wild-type animals from the dauer stage. By the above arguments, if ASI is the only sensory neuron that regulates dauer formation in unc-31 animals, those animals ought to be unable to recover from the dauer stage. We generated unc-31 dauer larvae by starvation and crowding, and examined their ability to recover and form adults after adding bacteria. 50/50 wildtype animals recovered from the dauer stage at 20 C in 24 hours, but only 5/122 unc- 31 animals recovered under the same conditions in a two week period. Thus unc-31 animals are extremely defective in recovery from the dauer stage. unc-31 animals will recover from the dauer stage when shifted to 15 C, but only slowly - half the dauers recovered in 5 days at 15 C. The recovery at 15 C might be caused by temperature-sensitivity of e928, or it might reflect the intrinsic temperature sensitivity of dauer formation noted in the study of other mutants.