Worm Breeder's Gazette 10(3): 92
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
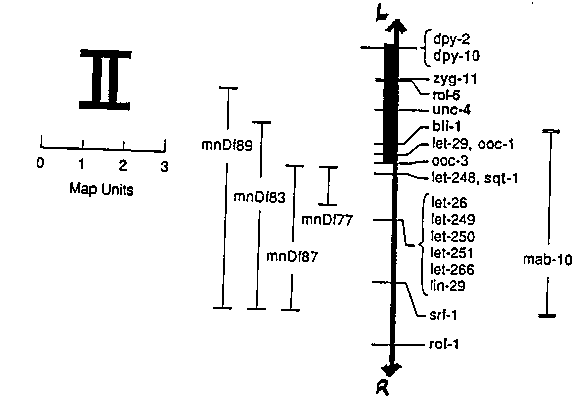
The recessive mutation mab-10(e1248) II was identified by J. Hodgkin in his screen for male-infertile mutants (Genetics 103:43-64, 1983). mab-10(e1248) males have rather subtle bursal defects, are completely infertile, and have been reported to occasionally have supernumerary molts. I have extended these observations and find that mab-10(e1248) causes a highly penetrant, male-specific retarded heterochronic defect in addition to affecting bursal development. Age-synchronized 248); 90) males and hermaphrodites were followed from the L4/adult molt at 20 C. Male animals were found to synchronously begin a supernumerary molt 18-20 hrs after L4 lethargis. Hermaphrodites of the same age (n=32) were carefully observed under DIC optics and showed no signs of a supernumerary molt. The male-specific supernumerary molt phenotype was found to be highly penetrant (>95%) at 16 C, 20 C, and 25 C. The male supernumerary molt appears to result from the production of a second adult cuticle. Young (presupernumerary molt) mab-10(e1248) adult males appear to have normal 'first' adult cuticles, as judged by the presence of adult lateral alae and wild-type staining with WGA and Ab117. The supernumerary cuticles also have adult lateral alae. To my knowledge, none of the known retarded heterochronic mutations (e.g., in lin-29, s pattern of adult supernumerary molts. The mab-10(e1248) supernumerary molt is not affected by passage through the dauer larvae stage. I have mapped mab-10(e1248) (scoring the supernumerary molt phenotype) to the right of the gene cluster on linkage group II, between unc-4 and rol-1 and near lin-29. (Male infertility has co- segregated with the supernumerary molt phenotype in these mapping experiments.) Interestingly, this places mab-10 in a region of the chromosome where the nearby known morphological mutations (in the genes lin-29, ifically affect the adult cuticle (see figure). mab-10(e1248) is complemented by lin-29 and the deficiencies mnDf87 and mnDf77, but not by the deficiencies mnDf89 or mnDf83. I have carefully observed the phenotype of mnDf89/mab-10(e1248) heterozygous males and hermaphrodites, and find the same pattern of male specific supernumerary molts as seen in mab- 10(e1248) homozygotes. This argues against the possibility that the male specificity of mab-10(e1248) results because this is a partial loss-of-function mutation and that a null mutation would affect both sexes. However, mnDf89/mab-10(e1248) males have significantly more abnormal bursae than mab-10(e1248) homozygotes, suggesting that mab-10( e1248) is not a null mutation. (mnDf89/+ males are wild-type.) Because the bursal defects (and male infertility) of mab-10(e1248) are observed before the supernumerary molt, the relationship between these phenotypes is unclear. It is also not obvious why a heterochronic defect would appear to be so sex-specific. In this regard, it is interesting to note that neither retarded nor precocious heterochronic mutations affect the timing of expression of male- specific bursal surface markers (Link, Ehrenfels, and Wood, Development 103:485-495, 1988). Perhaps some portion of male or bursal development involves an independent temporal program. [See Figure 1]