Worm Breeder's Gazette 10(3): 29
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
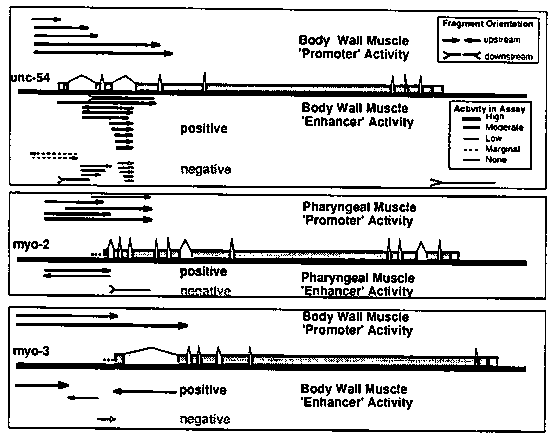
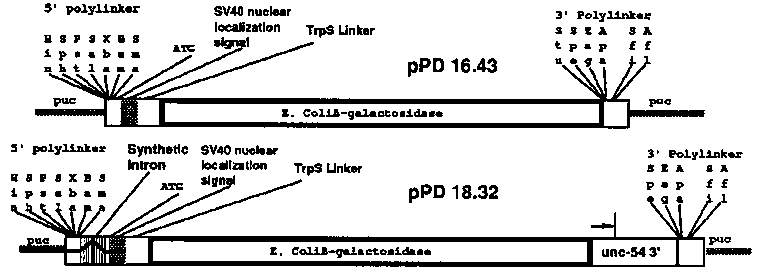
We have been using -galactosidase fusion vectors to define regions of the myosin genes that determine tissue specificity. The canonical fusion vector is shown below [16.43]. Our results with deletion mapping of the unc-54 gene (see last gazette) suggested that sequences at the 3' end of the gene, as well as the presence of at least one intron near the 5' end were important for gene expression. Thus we have constructed a more specific expression vector [18.32] which contains a synthetic intron sequence upstream of the -galactosidase coding sequence and the 3' region of the unc-54 gene downstream of B- gal. These vectors have been used for both transcriptional and translational fusions, and can also be used for chimeric intron constructs (splice fusions). [See Figure 1] A wide variety of -galactosidase fusion plasmids were constructed and tested either by creating and then staining transformed lines, or by an 'F1 Stain' protocol, i.e. injecting the DNA into oocytes of 5- 10 wild type animals and then staining the first generation. The latter procedure is surprisingly efficient, generating up to 10 stained progeny per injected adult; The stained F1 animals are often mosaics, with the extent of staining varying between one cell and the whole tissue. Thus a single set of 5-10 parents injected with a myosin- gal fusion generates 200-1000 labeled cells. To date we have not seen any difference in tissue specificity between transformed lines and 'F1 Stain' protocols; for routine characterization the 'F1 Stain' protocol is a rapid way of screening a large number of independent expression events. We define a 'tissue specific promoter' as any region that can be inserted into the upstream polylinker in 18.32 and lead to tissue specific expression. Segments upstream of the unc-54 and myo-3 genes ( body wall muscle myosins) and the myo-2 gene (pharyngeal myosin) have been inserted into 18.32 to make transcriptional fusions, and each construct leads to expression just in the expected tissue: Body wall muscle for unc-54 and myo-3, pharyngeal muscle for myo-2. Translational fusions with the first 2-5 exons of each of the three genes have also been constructed and these seem likewise to function only in the expected tissue. These translational fusions are each somewhat more active than their transcriptional fusion counterparts. The promoter fusions have been used to assay for 'tissue specific enhancer' function as follows: We start with a myo2- gal fusion construct that is only expressed in pharyngeal muscle, and insert segments from the other genes into various positions. We can then look for added expression in non-pharyngeal tissue. The best characterized enhancer to date is a sequence in the third intron of unc-54. This segment can be placed in either orientation upstream of the myo2- gal fusion and the resulting constructs exhibit strong -gal expression in body wall muscle as well as pharyngeal muscle. The segment also induces body wall muscle expression when placed 'downstream' of the myo-2 promoter region [of course in a 10kb. circular construct, 1kb downstream is equivalent to 9kb upstream ! ! ]. The effect of the enhancer is striking: without the enhancer the myo2- gal fusion is only expressed in the pharynx (>500 positively stained animals examined). With the enhancer present, >90% of the positively stained animals have strong body wall staining in addition to the pharyngeal staining. No enhancement of pharyngeal staining is seen. The left edge of this enhancer has been defined to within 25nt by deletion analysis and corresponds to an imperfect inverted repeat in the DNA sequence [nt 1972-2010]. The right edge of the enhancer has not yet been precisely defined, but maps within 170nt of the inverted repeat region. As in most eukaryotic systems, the distinction between promoter and enhancer is a subtle one. The unc-54 promoter region acts very weakly as an enhancer when placed upstream of the myo2- gal fusion; likewise neither the enhancer nor the promoter region is absolutely necessary for unc-54 gene expression (see last gazette): either the promoter ( all upstream sequences) or the enhancer can be deleted with retention of gene activity. Thus the requirement for the enhancer for gene expression is only manifest when deletions are made in the promoter region. We reported in the last gazette the apparently non-specific requirement for an intron near the 5' end for expression. This phenomenon seems to be separate from the enhancer function, i.e. introns without any enhancer activity are sufficient to satisfy the requirement. A survey of segments around the myo-3 gene has identified three segments with enhancer activity. One of these segments contains an inverted repeat sequence similar to the unc-54 enhancer. Pharyngeal 'enhancer' activity around the myo2 locus has been studied by a similar assay: myo2 segments placed upstream of the myo3- gal translational fusion can induce pharyngeal expression. We have begun a more general survey of worm sequences for enhancer activity. Briefly segments of C. elegans DNA are inserted into the myo2- gal fusion construct upstream of the myo2 promoter. These clones are then injected alone or in pools of 24 followed by staining of the F1. This protocol should pick up segments that can 'enhance' the myo2 promoter in tissues other than the pharynx. To date we have screened ~75k.b. of sequences and found at least four 'enhancer like' elements. In each case the expression of the construct is extended to muscles outside the pharynx, but evidently not to non-mesodermal tissue. There are qualitative differences between the 'enhanced' patterns, with one of the segments yielding particularly strong expression in the intestinal and sphincter muscles. The limitation of expression to muscles in these random constructs presumably results from a restriction to muscle expression of some element (e.g. the promoter) in the myo2- gal 'enhancer trap' vector: It is thus quite possible that the enhancers we have found in random C. elegans DNA segments might actually work in non-muscle tissues when tested on a more general promoter element. [See Figure 2]