Worm Breeder's Gazette 10(3): 23
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
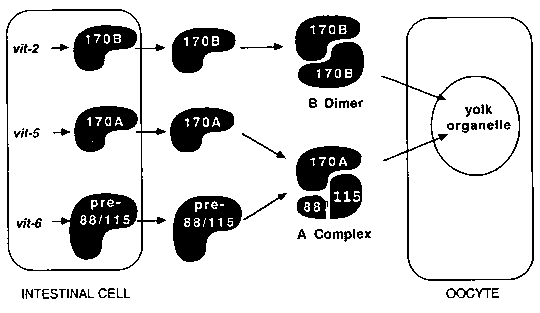
A number of observations on yolk proteins suggest that their primary biological function is lipid transport and sorting. It has been known for some time that the yolk proteins of vertebrates form dimeric lipoprotein complexes containing 15-20% lipid. But there was no obvious parallel between the vertebrate yolk polypeptides - two lipovitellins and phosvitin - and the four C. elegans yolk proteins - yp170A, yp170B, yp115, and yp88. Recently, Nardelli et al. (1) compared the vitellogenin genes from frog and chicken with the vit-5 gene from C. elegans and found substantial amino acid sequence similarities in many regions of the proteins. The exception is the phosvitin domain, which is not present in the nematode vitellogenins. Work in my laboratory has now produced a model of native yolk protein structures in C. elegans that confirms the conservation of higher-order yolk lipoprotein structure over the evolutionary interval separating nematodes from vertebrates. There are at least two distinct lipoprotein complexes in C. elegans. Both resemble the vertebrate complexes in that they are fundamentally dimeric, have relative molecular masses of about 480,000, and carry about 20% lipid, a mixture of phospholipids, triglycerides, and unidentified minor lipids. There is a twist, though. One of the C. elegans lipoproteins seems to be a simple homodimer of yp170B (the B dimer), resembling the vitellogenin dimers of vertebrates. But the other complex is apparently composed of yp170A, yp115, and yp88 (the A complex). It still has the properties of a dimer, suggesting that yp115 and yp88 together form a single monomeric functional unit equivalent to either of the yp170s. This is consistent with their origin in a single primary translation product, encoded by the vit-6 gene. The diagram below summarizes the model in the general context of vitellogenesis. [See Figure 1] It should be emphasized that this is a model. It accounts for the physical properties of yolk protein particles and for the results of immunoaffinity binding experiments, postulating only two mutually exclusive sets of protein-protein interactions. It is not possible, however, to exclude rigorously the existence of other combinations of the yolk polypeptides - a yp170A homodimer, for example, or a vit-6 'dimer' containing two each of yp115 and yp88. The sequence similarities detected by Nardelli et al. suggest that functional constraints have dictated the conservation of certain yolk protein characteristics in the evolution of birds and amphibians from invertebrates: the binding of lipids, the pairing of yolk lipoproteins in dimeric complexes, and the positions of cysteine residues. The binding of lipid appears to be a relatively autonomous function of the monomeric units. It's unclear, then, what advantage is gained by association of monomeric lipoproteins in dimers, but part of the reason for cysteine conservation seems to be the formation of intermolecular disulfides between the two halves of the dimers. There are indications of cross-linking between yp170A and yp88 in the A complex, and between the two yp170B polypeptides in the B dimer. Other cysteines are apparently involved in intramolecular disulfides. The association of yolk proteins with lipids suggests that lipid transport may be the primary function of yolk proteins. Because early embryogenesis in C. elegans and many other organisms proceeds essentially by subdivision of the zygote into progressively smaller membrane-bounded compartments, fairly large amounts of lipid are required for new membrane assembly. It may be advantageous to provide a maternally synthesized reserve of lipids so that everything doesn't have to be made up from simple precursors in the embryo. Such a lipid transport and storage role could explain the apparent variation of yolk protein utilization rates in C. elegans embryos (2). Yolk protein antigens seem to disappear from the anterior of the embryo, where cells are small and densely packed (hence, have a high proportion of membrane surface to volume) while they are retained in the intestine (large cells, with lower surface to volume ratio). The problems of moving lipids around in cells and organisms are universal. In fact, vitellogenins seem to be structurally related to at least one human serum apolipoprotein. Baker (3) has detected sequence similarities between vertebrate and nematode vitellogenins and human apolipoprotein B-100, the apoprotein of low-density lipoprotein (LDL). These homologies are sufficiently strong to imply that the human protein evolved, at least in part, from yolk proteins. Further, like LDL, vitellogenins are synthesized by the liver or intestine, bind to a specific receptor, and are endocytosed by the target cell. The big difference between yolk lipoproteins and serum lipoproteins may be that yolk proteins, with their bound lipid, are temporarily sequestered in yolk organelles after endocytosis, and are degraded only later, when the lipid reserves are needed.