Worm Breeder's Gazette 10(3): 162
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
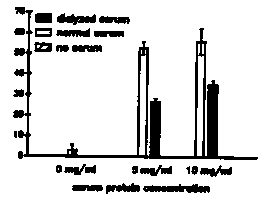
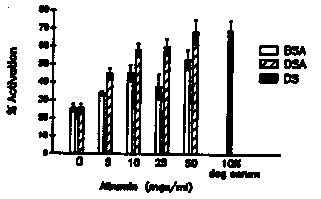
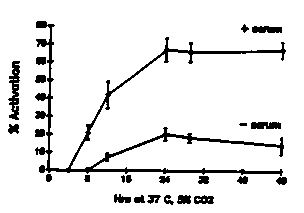
The developmentally arrested , free living third stage (L3) larva of hookworms resumes development upon entry into the host. This activation to the parasitic L3 occurs in response to a signal encountered in the host and may involve the reinitiation of developmental pathways previously arrested in the free living L3. Little is known about this activation signal in hookworms, due at least partially to the lack of a reliable in vitro assay for the resumption of development. The resumption of feeding, suspended in the free living L3, would be a logical prerequisite for the development to the parasitic L3 and beyond. With this in mind, we have developed an assay for the resumption of feeding in hookworm L3's, and, using this assay, have begun to characterize the host signals that activate L3's of the dog hookworm Ancylostoma caninum. This assay could also be utilized to determine the resumption of feeding in the recovering dauer larva. The assay: Hookworm L3's are recovered from 7-10 day charcoal-feces cultures, washed, and axenized by overnight incubation in sterile distilled water containing antibiotics (PST '100': penicillin 100 U/ml; streptomycin 100 g/ml; tetracycline 100 g/ml). Axenized larvae are washed in sterile dH2O, dilution-counted, and incubated in sterile 96 well microtiter plates, with approximately 250 L3's per well in a final volume of 0.1 ml RPMI 1640 (+ 25mM HEPES, pH 7.2, and PS '100') alone or supplemented with 10% (v/v) normal dog serum. Plates are incubated at 37 C in 5% C02 for the times indicated. After incubation, feeding larvae are labeled by the method of Clokey and Jacobson [Mech. Ageing. Dev. 35:79 1986] with modifications. Larvae are incubated with fluorescein isothiocyanate-labeled BSA (FITC-BSA) at 5 mg/ml ( final volume = 0.2 ml) for 1-2 h at 37 C, 5% C02, then washed in 15 ml 0.9% NaCl. Larvae that have ingested the labeled albumin (i.e. activated) are counted using a Zeiss standard microscope equipped with epi-fluorescence illumination (excitation 450-490 nm, barrier 515-565 nm) and expressed as a percentage of the total L3's counted. A minimum of 50 L3's are counted from each of 3 replicates. The results: Populations of A. caninum L3's begin feeding between 6 and 8 h after incubation in the presence of serum, as indicated by the appearance of fluorescence in the esophageal and intestinal lumina under UV illumination. Feeding peaks by 24 h, with maximum activation levels of 70-90%, and remain at this level through 48 h (fig. 1). Larvae incubated in the absence of serum activate at much lower levels (<20 %). Additionally, 48 h-activated L3's tend to curl into tight loops, whereas larvae in wells lacking serum do not, indicating that an activation-associated behavioral change has occurred also. The serum dependence suggests that the activation signal is a component of serum. Ultrafiltrated serum shows stimulatory activity in both the <10 kD and >30 kD fractions (fig. 4), and dialysis of whole serum reduces the activity by almost half (fig. 2), suggesting that a small (<8000 MW) molecule is stimulatory. However, the ability of the >10 kD and >30 kd fractions to initiate feeding indicates that a larger MW factor is also stimulatory. Both dog serum albumin (10 mg/ml) and bovine serum albumin (50 mg/ml) activate L3's to near maximum levels (fig. 3). Other proteins, such as alpha- and gamma- globulins, ovalbumin, and tryptone, fail to activate L3's at concentrations to 100 mg/ml, indicating that the stimulatory effect of albumin on feeding is not caused by the presence of protein (i.e. 'food') alone. Dialysis of BSA has no effect on its stimulatory ability, indicating that the activity is probably not due to a loosely bound ligand. These data suggest that there are two components of the activation signal, a small MW factor and albumin, although other large molecules not tested may also be stimulatory. Further characterization of both the large and small fractions is required. [See Figures 1-4]