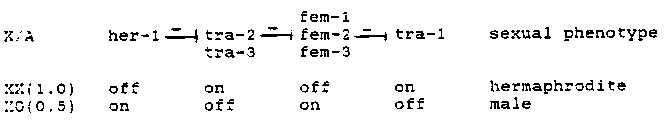
Worm Breeder's Gazette 10(3): 15
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
The fem-3 gene is required for specification of the male fate. In hermaphrodites, it specifies spermatogenesis and is contributed maternally to influence progeny sex; in males, it specifies male development in all tissues. Previously we have shown that fem-3 RNA is present in adult hermaphrodites, but that it is absent from adult hermaphrodites that lack a germ line [either SS104(bn2) or glp-1(q224) 1 (Rosenquist and Kimble, 1988). We interpret this to mean that fem-3 RNA is limited to the germ line in hermaphrodites. We have now used SS104 and glp-1 mutants to examine the presence of fem-3 RNA in adult males with and without a germline. In contrast to hermaphrodites, we find that fem-3 RNA is detectable in XO males whether or not they have a germline. We interpret this to mean that fem-3 RNA is present in XO somatic tissues. To examine the molecular basis of the Hodgkin model in which the sex determination genes are proposed to act in a cascade of negative regulation (see below), we are in the process of testing double mutants for the presence of fem-3 RNA in XX animals masculinized by a tra(lf) mutation or for the absence of fem-3 RNA in XO animals feminized by a fem(lf), her(lf), or tra(gf) mutation. Preliminary data indicate that XX pseudomales of genotype bn2;tra-3( e1107)(m-z-) do contain somatic fem-3 RNA in the XX soma. In addition, we have shown by primer extension experiments that fem-3 RNA is trans- spliced to the 22bp leader RNA described by Krause and Hirsh.