Worm Breeder's Gazette 10(3): 130
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
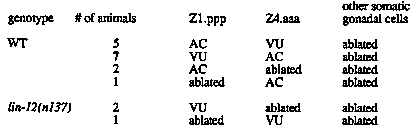
In wild type hermaphrodites, the fates of the somatic gonadal cells Z1.ppp and Z4.aaa are naturally variable: with equal probability, one becomes the anchor cell (AC) and one becomes a ventral uterine precursor cell (VU) (Kimble and Hirsh, 1979); cell-cell interactions govern this choice of cell fates (Kimble, 1981). Genetic and anatomical characterization of lin-12 mutants indicated that lin-12 activity is necessary and sufficient for the specification of the VU fate (Greenwald et al., 1983). We have been investigating the role of lin-12 in VU fate specification by a combination of laser ablation experiments and genetic mosaic analysis. The goal of these experiments is to ascertain if lin-12 function is required in the presumptive VU cell itself (i.e., lin-12 function is cell autonomous) or in another cell, such as the presumptive AC or in another gonadal cell (i.e., lin-12 function is cell nonautonomous). This question is of particular interest because lin-12 appears to encode an integral membrane protein, which could conceivably function as a receptor/signal transducer (cell autonomous) or as a signal (cell nonautonomous) (Yochem et al., 1988). From the work summarized below, we conclude that lin-12 function is likely to be cell autonomous in the AC/VU decision, which is inconsistent with models in which lin-12 protein is proposed to serve as an intercellular signal. We extended the observations of Kimble (1981), who demonstrated that an isolated Z1.ppp or Z4.aaa invariably assumes the AC fate, suggesting that the AC fate requires no cell interactions for its establishment. We ablated all somatic gonadal cells except Z1.ppp and Z4.aaa, and showed that one cell becomes an AC and the other becomes a VU. This result suggests that the VU fate depends only on the presence of an AC and does not require interactions with any other gonadal cell(s) (Table 1). We also screened by Nomarski microscopy for genetic mosaics in which either Z1 or Z4 was genotypically lin-12(0) in an otherwise lin-12(+) background, using the strain qDp3; 65) 51) 41). [qDp3 carries wild type alleles of these three genes (Austin and Kimble, 1987); ncl-1 is recessive and results in abnormal nucleolar morphology (E. Hedgecock, personal communication).] We obtained 12 such mosaics: in each of the 12 cases, only a single functional AC was formed and this AC was genotypically lin-12(0) (Table 2). This result suggests that lin-12 expression is not required in the AC for correct VU specification. In addition, it is interesting to note the apparent bias towards the AC fate of the lin-12(0) cell [as opposed to the lin-12(+) cell] in our mosaics. In wild type, Z1.ppp and Z4.aaa have an equal probability of becoming the AC; thus, we might have expected only 6 out of our 12 mosaics to have a single lin-12(0) AC. We are currently investigating this point further. The laser ablation and genetic mosaic data together imply a cell autonomous role for lin-12 activity in the AC/VU decision: the laser experiments demonstrate that only Z1.ppp and Z4.aaa interact to establish the VU fate, and the mosaic experiments demonstrate that lin- 12 does not function in the AC to establish the VU fate. Additional evidence supporting this conclusion derives from laser ablation experiments in lin-12(d) hermaphrodites. In lin-12(d) hermaphrodites both Z1.ppp and Z4.aaa become VUs. We have found that in lin-12(d) hermaphrodites, an isolated Z1.ppp or Z4.aaa becomes a VU, not an AC ( Table 1). We are currently testing the cell autonomy of lin-12 function in other decisions apparently controlled by lin-12 by isolating additional mosaics. Ultimately, we would like to express lin-12 specifically in defined cells in order to test the conclusion of cell autonomy based on genetic and developmental studies. [See Figures 1 & 2]
