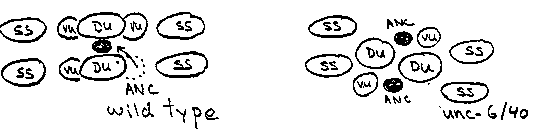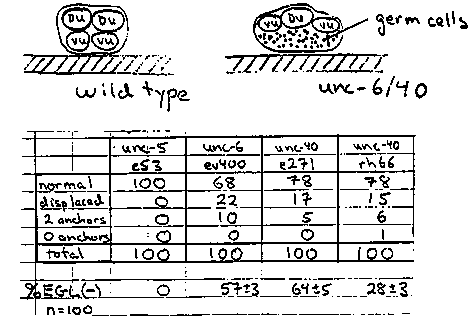
Worm Breeder's Gazette 10(3): 122
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
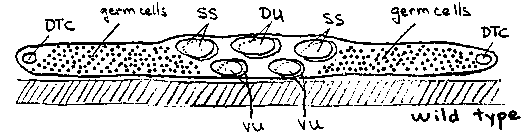
The embryonic gonad comprises four cells pressed against the ventral body wall (Fig. 1). Cell divisions in the first larval stage create 2 distal tip cells (DTC), 4 sheath/spermathecal (SS), 2 dorsal uterine (DU), and 4 ventral uterine (VU) precursors plus mitotic germ cells ( Fig. 2). The somatic cells, excepting the distal tip cells, coalesce during the second larval stage, displacing all germ cells from the central region of the gonad. The 6 uterine cells form a central prism with the 4 VU cells contacting the body wall and 2 DU cells facing the body cavity. The fates of the four ventral uterine cells are regulated by interactions between the uterine cells or their immediate descendants. Most strikingly, either Z1.ppp or Z4.aaa, becomes the unique anchor cell, the primary fate for this pair of equivalent cells. The committed anchor cell moves to the center of the ventral uterus and induces vulval development (Fig. 3). Division axes and early movements of gonadal cells may be determined by affinities between gonadal cells and by affinities of specific somatic cells, e.g., distal tip cells, for the body wall. Here we report that unc-6 and unc-40 mutations disturb the arrangement of the hermaphrodite gonad primordium perhaps by disrupting adhesion of VU cells, including the anchor cell, to the epidermis. unc-6 and unc-40 hermaphrodites are often absolutely defective in egg-laying (see Fig. 4). We examined late third stage larvae by DIC microscopy. In about 20% of the animals, the anchor cell was displaced laterally or dorsally in uterus and failed to connect to the developing vulva. Such animals were incapable of laying eggs as adults. In another 5-10% of the larvae, 2 anchor cells were present, often widely separated on the uterine circumference. We examined early third stage larvae to learn how these anchor cell displacements or duplications arise. The arrangement of the somatic primordium was disturbed in many individuals. Most cell-cell contacts within the somatic primordium were preserved in these mutants. In particular, SS- DU, DU-DU, and DU-VU contacts appeared to be normal. The relative positions of the 4 VU cells, however, or their relation to the body wall, were frequently abnormal. In particular, left and right VU cells sometimes failed entirely to meet each other. (Two anchor cells were formed in these animals suggesting that the regulative interaction between Z1.ppp and Z4.aaa mediated by the lin-12 protein requires close proximity or direct cell contact.) In many individuals, the VU cells were displaced from the epidermis by germ cells (Fig. 4) . We have not yet examined younger larvae to learn whether these abnormal cell arrangements arise from altered L1 division axes or failed L2 cell movements. A simple model to account for these observations is that VU cells adhere to the underlying ventral epidermis. This adhesion brings or holds the left and right VU cells together at the ventral midline, establishes the dorsal/ventral orientation of the uterus relative to the body wall, and enables the uterine cells to squeeze germ cells out of the central region of the gonad. In unc-6 and unc-40 mutants, this adhesion is defective and the uterine cells can be pried from the body wall by intruding germ cell. This model is consistent with observations that these genes may guide other ventralward cell movements on the body wall.