Worm Breeder's Gazette 10(3): 116
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
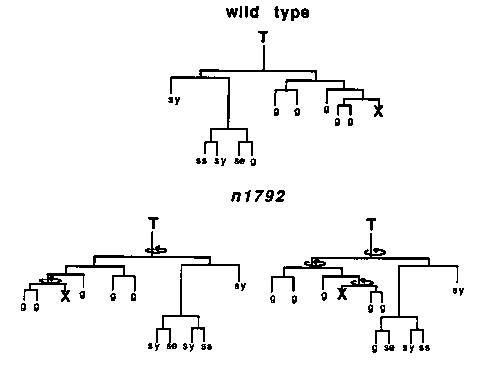
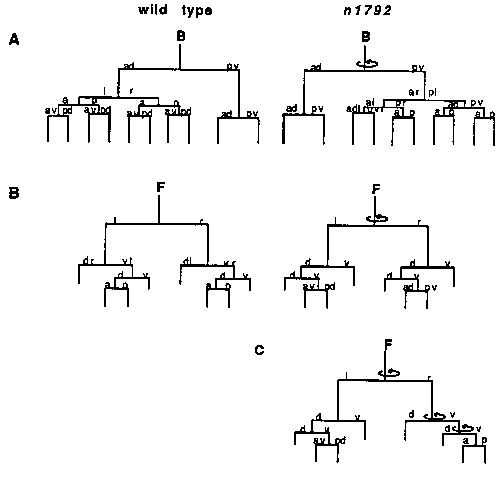
During C. elegans development most cell divisions are asymmetric in that the two sister cells generated differ in their cell fates. Mutations that either cause normally asymmetric cell divisions to become symmetric or that reverse the polarity of the asymmetry may identify genes that control asymmetric cell divisions. Three such genes have been previously identified: lin-11, ns in each of these genes affect cell fates within the vulval equivalence group. Mutations in lin-11 primarily cause asymmetric cell divisions to become symmetric (see Freyd et al, this newsletter), while mutations in lin-17 and lin-18 cause both loss of asymmetry and reversal of the polarity of the asymmetry. In addition, lin-17 affects the lineages of T, Z1 and Z4 in hermaphrodites and males, and P10.p, P11.p and B in only males. We recently isolated a new mutation that affects certain asymmetric cell divisions. This mutation, n1792, was identified because it results in deformed male tails. In the dissecting microscope all n1792 males appear to have club-like tails with reduced or missing fans, although the severity of this defect varies. Most n1792 hermaphrodites appear wild-type in the dissecting microscope, although some are slightly Egl. The tail defect of n1792 males led us to determine the lineages of the male-specific blast cells B and F. We followed the B lineage of seven n1792 males through the L2 molt. One had a wild-type lineage, five had a lineage similar to the one shown in Figure 1A and in one the B cell divided symmetrically generating four hypodermal-like cells. We determined the F lineage of six n1792 males. One had a wild-type lineage, three had a F lineage similar to the one shown in Figure 1B and in one the F cell divided symmetrically generating eight cells. Shown in Figure 1C is an animal that appeared to reverse polarity at multiple points in the F lineage (points of polarity reversal are indicated by the circular arrows). It is interesting to note that the B lineage has an anterior-posterior asymmetry while the F lineage has a dorsal-ventral asymmetry, and both are reversed in n1792 males. We have also determined the lineages of the male-specific blast cells Y and U in n1792 males. We determined the complete Y lineage in one n1792 male and did a partial lineage of two others. All appeared wild-type. In addition, the first division of Y is asymmetric in wildtype males, so that Y.a is much smaller than Y.p. We have observed several first divisions of the Y cell in n1792 males and all appear wild-type. We determined the U lineage of seven n1792 males. In wildtype males the U lineage is symmetric until the last division, in which either one or both of the cells U.l/ra divides. In three n1792 males the U lineage was wild-type, in two animals U.1/rp divided (polarity reversal) and in two animals neither U.1/ra nor U.1/rp divided (symmetric). Thus it appears the U lineage can also be reversed in n1792 males. However, since the asymmetric division of the U lineage in wild-type males is variable, and in the deformed tails of n1792 males, the cells are in abnormal positions, it is not clear whether the effect of n1792 on the U lineage is direct or is due to the abnormal environment. Since all the defects of n1792 animals we have found so far are in the posterior region of the male, we determined the T lineage in two n1792 males to the mid L2 stage. Both were reversed in polarity. We then determined the complete T lineages in two n1792 hermaphrodites ( Figure 2). Both mutant lineages have multiple points of polarity reversal. Since the T lineage gives rise to many easily distinguishable cell types, determining the T lineage of many n1792 hermaphrodites will allow us to examine the phenomenon of multiple polarity reversals in more detail. Two of the cells produced by the T lineage, T.paa and T.pap, are the phasmid socket cells PHso1 and PHso2. The phasmids are sensory structures located in the tail consisting of two pairs of neurons that are open to the environment via the phasmid sockets. If wild-type animals are soaked in the fluorescent dye FITC, the phasmids fill with FITC, as do the amphids (sensory structures located in the head), and can be seen in the fluorescence microscope. Previously, Ed Hedegcock showed that mutations in lin-17 cause the T cell to divide symmetrically, so that the phasmid socket cells are not made. He showed that this defect blocks FITC filling of the phasmids. With this in mind, we soaked n1792 hermaphrodites in FITC and observed them in the fluorescence microscope. Of 59 animals examined, none of the phasmids filled with FITC, whereas the amphids filled in all 59 animals. To determine whether PHso1 and PHso2 or the phasmid neurons are defective, we examined FITC filling in early L1 n1792 animals . In early L1 wildtype animals, the T cell serves as the phasmid socket before it divides Since the phasmids of 49/50 early L1 n1792 animals filled with FITC, it appears that PHso1 and PHso2 (products of the T lineage) are not functioning in older n1792 animals . There are two possible explanations for the lack of function. Since we cannot tell by appearance whether the cells generated are actually PHso1 and PHso2, it is possible that PHso1 and PHso2 are not made. Alternatively, the abnormal position of PHso1 and PHso2 in n1792 animals may block their function. In addition, we have determined the lineages of the following cells in n1792 animals and all were wild-type: H1, H2, V1-V6 to completion in two animals and through the L3 stage in two others, QL in two animals and QR in one animal. We have also examined the hermaphrodite vulval cells in 14 L3 and L4 animals and all appeared wild-type. While examining L3 and L4 males under Nomarski optics we noticed that P12.p had expressed a P11.p-like fate in several n1792 males . In fact 7/36 n1792 males showed this transformation, including one male in which we determined the lineages of P11 and P12. In this male P12.aap divided in the L3 (normally P11.aap but not P12.aap divides), suggesting the transformation is at the level of the P cells themselves. Thus in contrast to the effect n1792 has on the asymmetric cell divisions described above, here n1792 has an effect on cell fate . Since we know that the cells P11 and P12 interact to choose their cell fates, and they are affected by n1792, it is possible that the asymmetric cell divisions affected by n1792 also are involved in intercellular interactions . It is interesting that all the cells affected by n1792 are in close proximity. Furthermore, at least one of the defects may involve intercellular interactions. Perhaps cell-cell communication is involved in the generation of asymmetry in these cell lineages and n1792 mutants are defective in this common signalling system. [See Figures 1 & 2]