Worm Breeder's Gazette 10(3): 110
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
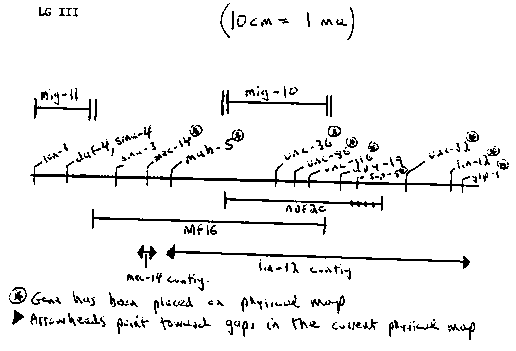
I am continuing studies that I initiated in Boulder of several cell migration mutants [Manser and Wood, manuscript in preparation; also, e. g., WBG 9(3), p.91 and 9(2), p.63, 1986]. To investigate how the corresponding genes are involved in cell movement, I would like to isolate and use DNA clones and perform mosaic analyses. At present, I am focusing on the mig-10(ct41)III and mig-11(ct78)III genes. In most mig-10(ct41) embryos, neurons ALM, CAN, and HSN migrate only partway toward their normal destinations (sometimes the ccL mother cells do likewise). In mig-11(ct78) embryos, CAN migration is rendered partially defective with high penetrance, while ALM and HSN migrations are affected at very low penetrance. Previously obtained genetic map data place both loci within the lon-1 to unc-32 cluster on LGIII, a region favorable for mosaic analysis (qDp3, unc-36; see Austin and Kimble Cell 51: 589-599, 1987), and one that is partially covered by the current physical map (lin-12, ibly ced-4, contigs; see Coulson et.al., this issue). However, more precise map positions are required to construct strains for mosaic analyses and to determine whether the current physical map will be of use for cloning. Thus I have been doing some additional mapping. I have found that mig-10(ct41)III lies within the interval covered by both nDf16 and nDf20. mig-10 thus maps within the mab-5 to dpy-19 interval (see Figure), which is completely spanned by the lin-12 contig. More precise positioning of mig-10 within this interval should allow me to select a small set of cosmids for use in transformation rescue experiments (Hope et.al., WBG 10(2) pp.97-98, 1988) or, if rescue cannot be achieved, to devise alternative cloning strategies that make efficient use of the physical map (e.g., searches for polymorphisms in mutants). Thus I am currently mapping mig-10 relative to mab-5, unc-86, dpy-19 have been placed on the current physical map). Results from the deficiency mapping experiments also suggest that the ct41 allele is null or nearly null. Specifically, I examined several phenotypes of ct41/nDf20 animals ( adult viability, adult dissecting microscope phenotypes, and cell migration phenotypes as determined by Nomarski microscopic examination of L1's), and found none to be more severe than what is observed for ct41 homozygotes. Even if ct41 is not a true null, it should be possible to isolate such an allele in a complementation screen with ct41. I have positioned mig-11(ct78)III within the lon-1 to sma-3 interval in three-factor and deficiency (nDf16 and nDf20) mapping experiments ( see Figure; complementation tests with nDf16 indicate that mig-11 maps to its left). Because lon-1, genes between them (daf-4 and sma-4) have not yet been assigned to contigs, it is possible that mig-11 is absent from the current physical map. Nevertheless, I am currently mapping mig-11 relative to lon-1, sma-4, and sma-3 in anticipation that the physical and genetic maps will eventually become better correlated in this region. I also plan to continue my studies of the vab-8V gene (whose apparent null phenotype includes a highly penetrant CAN migration defect: Manser and Wood, op. cit.). The close proximity of vab-8V to myo-3V on the genetic map suggests that vab-8 may be contained in the myo-3 contig; if so, the physical map may prove useful for cloning. [See Figure 1]