Worm Breeder's Gazette 10(2): 87
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
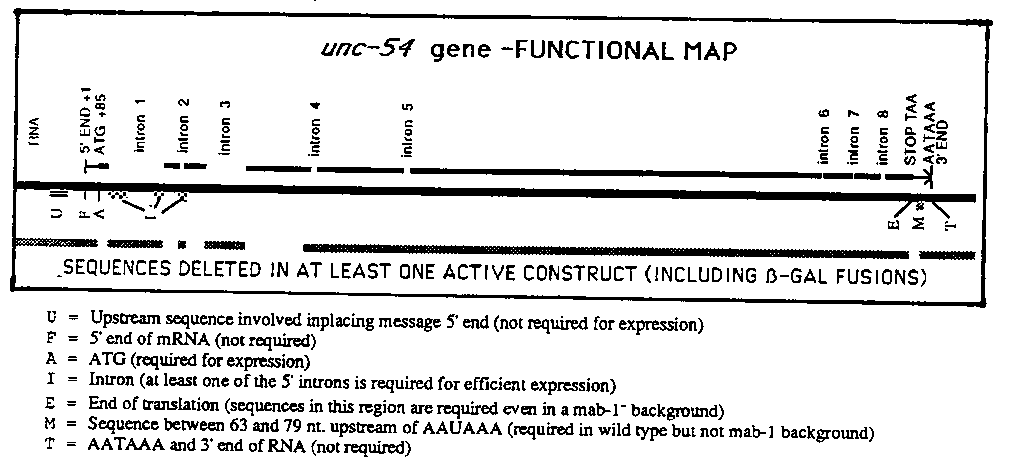
In the last gazette, we reported the beginnings of a deletion analysis of the unc-54 gene. We have since extended this analysis. To date, several surprising results have emerged. Methods: Plasmid clones carrying modified or deleted copies of the unc-54 gene are constructed. These plasmids are injected into the oocytes of unc-54(e190delta) animals. If the particular plasmid is active, then a high fraction (5-25%) of the injected oocytes give rise to animals that are rescued in the sense that their movement is greatly improved over e190 or mock-injected control animals (also the rescued animals are often egl+). A small fraction (3-20%) of the rescued animals give rise to heritable transformed lines. In order to assess tissue specificity, these lines are stained with a mixture of D. Miller's monoclonal antibodies. Upstream Sequence Requirements: Essentially there are no sequences upstream of the unc-54 gene that are required for tissue specific expression. This result is described in some detail in our article in the last gazette. Since then the 'ultimate' upstream deletion has been engineered, missing all upstream sequences to +79 from the site of transcriptional initiation (6bp upstream of the AUG). This construct is completely active for unc-54 function. Transformed lines carrying the +79 deletion give the wild type pattern of tissue specific expression. Deletion of the next 21 bases (including the AUG) results in an inactive gene. Since each of the deletion mutants assayed so far had substituted plasmid sequences for the deleted sequences at the 5' end of the gene, we decided to replace the 5' flanking sequences with DNA flanking another nematode gene. We chose the myo-2 gene, which encodes a myosin isoform made only in pharyngeal muscle (unc-54 and myo-3 are expressed in body wall type muscles). Therefore we substituted a region of 1660 bp from upstream of the myo-2 gene for the sequences upstream of the unc-54 message (the fusion site was at +50 in the unc- 54 message). This construct is active in rescuing the paralyzed phenotype of unc-54(e190). This result further supports the hypothesis that sequences inside of the unc-54 gene are sufficient for its expression in body wall muscle. When a transformed line carrying the myo2:unc-54 fusion was stained with the antibody mixture described above, it was found that unc-54 protein (but not myo-3 protein) was present in the pharyngeal muscles as well as body wall muscles of the transformed animals. This result shows that the observed differential localization of the myosin isoforms to the different muscle types is not a result of protein stability or differential detection. Interestingly, the unc-54 myosin seems organized into radial filaments when present in pharyngeal muscle. Intron Requirements: Intron sequences near the start of the message are apparently required for efficient expression of the gene. Deletion of the first four introns (using a synthetic cDNA clone provided by J. Karn) results in a plasmid (pUNK54delta1234) with almost no activity in transformation assays. By performing a very large number of injections few partially rescued animals have been obtained with this plasmid, and one transformed line has been obtained. This line is very sick (e.g. usually sterile), exhibiting a variety of abnormalities. A variety of plasmids with deletions of different subsets of introns has been constructed by cutting and pasting between pUNK54 and pUNK54delta1234. Surprisingly all of these constructs so far tested are completely active. This includes the plasmids pUNK54delta1, pUNK54delta234 pUNK54delta134, and a plasmid 'pUNK54deltam1+234', in which 389 b internal to the 562 b intron #1 have been deleted as well as introns 2,3,and 4.) In the cases of pUNK54delta1 and pUNK54delta234, transformed lines have been obtained and exhibit wild type patterns of unc-54 tissue specificity. The full activity of the four 'subset' plasmids has several consequences in terms of any explanation for why the introns are required. First, the intron subset deletions were constructed in such a way that every segment of the inactive plasmid pUNK54delta1234 was subsequently recloned in generating at least one of the fully active construct. Thus the inactivity of pUNK54delta1234 cannot be the result of some spurious mutation either present in the original synthetic cDNA construct or introduced during cloning. Indeed these data indicate that no single sequence within the first four introns is absolutely required for efficient unc-54 expression. Rather the requirement for 'something' within introns 1-4 can be satisfied at minimum either by the presence of the very short intron #2 (at 38 b. this is the shortest intron in unc-54), or by the trimmed intron #1 present in pUNK54deltam1+234. Although it is possible that this 'something' is some sort of redundant enhancer or promoter element that is present in both of these segments, it seems quite possible that there is a mechanistic requirement for a splice near the 5' end of the long unc-54 message in order for the message to be expressed. Particularly puzzling if this were true would be the fact that even the pUNK54delta1234 plasmid still has four introns, and that the contiguous unspliced 5' region in pUNK54delta1234 is actually shorter than the longest contiguous region in the wild type unc-54 message ( exon #6). Thus one might propose that certain worm messages need a splice somewhere near their 5' end. Downstream Requirements: We have also undertaken a deletion analysis of the 3' end of the gene. The sequence landmarks in the 203 nt 3' untranslated region of the message are an AAUAAA sequence (present upstream of most eukaryotic cleavage and polyadenylation sites), and the presumed the 3' end of the RNA (35 b. downstream of the AAUAAA; suggested by sequencing of cDNA by A. MacLeoud, S. Brenner, and J. Karn). Surprisingly neither of these sequences is necessary for function of the gene. This is shown by the full activity, both in transient assays and in appropriate expression in transformed lines, of a clone deleting all of the 3' flanking region back to 63 b upstream of the AATAAA. This construct has no AATAAA for at least 3kb downstream from the gene; a second construct replacing plasmid flanking sequences in this deletion with the sequences normally 3' to the unc-54 gene is also active. There do seem to be some 'required' sequences within the 3' untranslated leader. This is shown by a set of deletions which remove the entire 3' non-transcribed region back to 79, 108, and 116 nt upstream of the AATAAA. These constructs are inactive, i.e. they do not rescue unc54(e190) animals. Thus some sequence between 63 and 79 nt upstream of the AATAAA appears to be involved in proper expression of the gene. This region lies in the center of various folded structures that can be drawn for the unc-54 3' leader. Rock Pulak and Phil Anderson reported in the last gazette that a genetically isolated deletion (r293) in the 3' region of the endogenous unc-54 gene behaves as an unc-54 null. It is tempting to speculate that the inability of the 79, 108, and 116 deletions to function is related to the inactivity of r293. Pulak and Anderson also isolated and characterized suppressors of the r293 allele (which they and J. Hodgkin have shown correspond to a set of morpho-mabs described in detail in the last Gazette by Hodgkin, Papp, and Ambros). At the suggestion of Anderson and Pulak, we tested whether the requirement for sequences between -63 and -79 from the AAUAAA could be suppressed by one of these mab mutants. This was done by injecting the -79 deletion into unc-54(e1092)mab-1(e1228) animals. The -79 deletion is indeed active in these animals; a transgenic line PD107 was derived, and the resulting transgenic locus loses the unc-54 suppressing activity when Mab-1+ function is present. The pattern of expression of unc-54 in PD107 (as well as in a number of the mab- rescued r293 lines) is normal, suggesting that the identified sequences required in the the 3' leader probably do not play a role in tissue specificity. Puzzle: Some sequence(s) within the unc-54 clone must be responsible for tissue specificity. Either these are present within the coding sequences of the first three exons, in a short 3' region (~100nt) yet unexplored or redundant information is present so that no single sequence is necessary for tissue specificity. [See Figure 1]