Worm Breeder's Gazette 10(2): 78
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
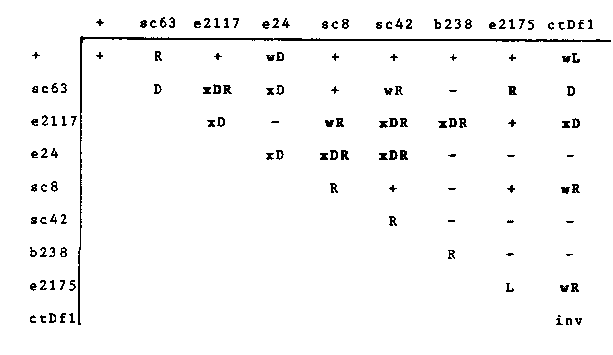
To understand the role of the sqt-3 gene in C. elegans cuticle development, we have constructed allelic combinations with known alleles of sqt-3(sc63, e24), rol-4(sc8, sc42, and b238), lon-3 (e2175) and ctDf1. To obtain progeny of type a/b, N2 males were crossed with a/a hermaphrodites to give a/+ males. These were mated to b/b hermaphrodites at 25 C to give a/b hermaphrodites among others. Suspected a/b worms were confirmed by allowing them to self at 25 C, and a resulting 1:2:1 segregation of a/a : a/b : b/b confirmed the phenotype. [See Figure 1] '-' indicates the cross was not performed. All rollers (R) are left rollers. Weak rollers(wR) were classified by their occasional twisting, particularly when prodded, and by their visibly twisted cuticle. Strong dumpy(xD) worms range from being slightly larger than eggs to about 50% N2 length, with a correspondingly wide range of mobilities. All alleles of rol-4 are recessive, as is sqt-3(e2117). The other two alleles of sqt-3(sc63 and e24) have dominant phenotypes ( roller and weak dumpy) as does ctDf1 which is a weak long (wL). Phenotypes not in bold face were obtained from Kusch and Edgar, Genetics 113: 621-639, 1986, or from Cox et al., Genetics 95: 317-339, 1980. If sqt-3 and rol-4 are different genes, the expected phenotype of the heteroallelic combinations would be either wild type or the phenotype of the dominant sqt-3 allele. An exception to this would arise in the case of intergenic noncomplementation, which has been seen for other cuticle defective mutations. Our results show that in many cases the combination of a sqt-3 allele and a rol-4 allele, both in heterozygous configurations, generates a doubly mutant phenotype. This phenotype is the combined display of what both alleles would show alone were they homozygous. For example, e2117/sc42 is a severe dumpy e2117/b238, e24/sc8, and e24/sc42. In contrast, sc63, the only true sqt allele (m/m = Dpy; m/+ = Rol) is suppressed by the two rol-4 alleles tested: sc63/sc8 is WT and sc63/sc42 is a weak roller. We have also determined that the phenotype of rol-4(sc8)/ctDf1 is a weak roller, suggesting that it is located within the ctDf1 region. If sc8 were not located in the deficiency, the expected phenotype would be a weak long, barring extragenic effects. This contradicts the assignment of rol-4 to the right of ctDf1 as is illustrated on the genetic map, 1987. We have no data that support the notion that sqt-3 and rol-4 are separate loci, although we have not yet determined the frequency of recombination between sqt-3 the assignment of these depend on sequence data of and rol-4 alleles. Ultimately, alleles to one or the mutant DNAs. Lon-3(e2175) may also be located in the ctDf1 region since e2175/ctDf1 is a weak roller, not WT like e2175/+. Lon-3(e2175) does not interact with any other allele tested, however, and is unlikely to be allelic with sqt-3 / rol-4.These complementation data allow us to predict the range of phenotypes that may be generated as additional mutations closely linked to sqt-3. We are beginning to generate new mutations that fail to complement sc63 using [32P] as the mutagen. Such new mutations are likely to have DNA alterations detectable by Southern blotting (Hoskins et al., WBG 10, No. 1). We will probe these mutant DNAs with the collagen genes which we have isolated from the myo-3 contig (Edwards, WBG 10, No. 1).