Worm Breeder's Gazette 10(2): 29
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
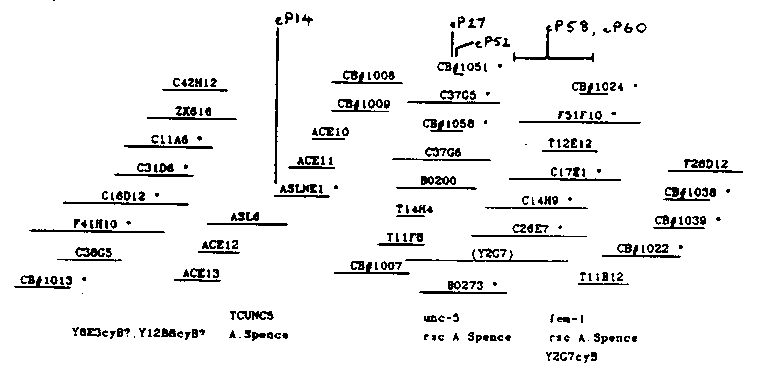
The fem-1 gene is essential for all aspects of male development and for spermatogenesis in the hermaphrodite in C. elegans (Doniach and Hodgkin. Dev. Biol. 106:223.1984). It is located 0.03 m.u. to the right of unc-5 on LGIV. The figure summarizes the present state of the contig which I described in the last WBG (10#1:33) and which now contains both unc-5 and fem-1.[See Figure 1] The orientation of the contig on the genetic map is established by two polymorphisms, eP14, and eP60, present in strain CB3724, which is congenic between N2 and N62 over the unc-5 - mor-2 interval, and which was constructed by T. Doniach. eP14, as I mentioned in the last WBG, maps to the left of unc-5, while eP60, detected by cosmid C26E7, maps to the right. [One of 16 recombinants in the unc-5-mor-2 interval separated eP60 from unc-5(+)]. Between these two polymorphisms are located several which cosegregate with various unc-5 alleles [eg. eP27 with unc-5(bx8), eP52 with unc-5(st1003)]. Cosmid C26E7, and the overlapping clone C17E1, detect a polymorphism, eP58, in the X-ray- induced allele fem-1(e2044). eP58 cosegregated with e2044 in 15 of 15 recombinants in the fem-1-mor-2 interval. A 15 kb fragment from the region of overlap between these two cosmids reveals not only eP58 but also polymorphisms in each of three [32P]-induced fem-1 alleles, e2365, e2365, and e2382. The same 15 kb probe detects two transcripts of about 3 kb and 2.7 kb in mixed populations of either N2 hermaphrodites or him-8 hermaphrodites and males. I am presently locating the fem-1 polymorphisms and these transcripts more accurately in the hope of identifying one or both transcripts as products of fem-1. Cosmid C17E1 is able to transform fem-1 XX females into self-fertile hermaphrodites. I injected females of the genotype fem-1(e2044)mor-2( e1125). e2044 being a putative null allele of fem-1, and mated them with fem-1(hc17ts) males raised at 15 C. The F1 were raised at 25 C, at which temperature the XX animals develop as females and the XO animals as sterile intersexes unless transformed to fem(+). I obtained five rescued F1,and from the progeny of three I was able to start lines homozygous for each of the two fem-1 alleles. The rescuing DNA is transmitted as an unstable duplication in each line. ( See the note by Hope et al. in this issue for a summary of the injection results.) I have not yet measured the fertility of the XX hermaphrodites in these lines, but it is lower than wild-type at 25 C At least some of the XO animals appear to be normal males, but I have not tested their fertility. I intend to define the extent of the fem- 1 locus more accurately by repeating the transformation experiments with smaller DNA fragments from the region in which the fem-1 polymorphisms reside.