Worm Breeder's Gazette 10(2): 150
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
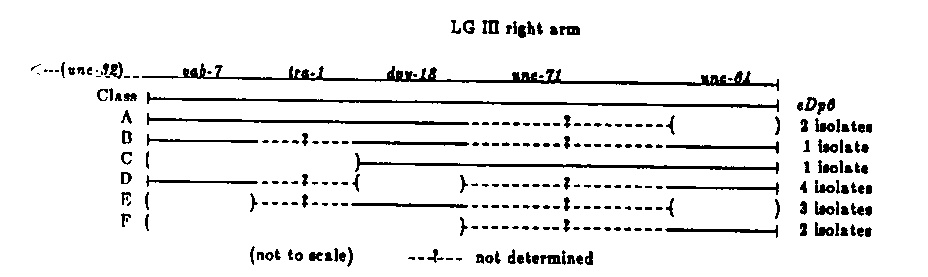
The free duplication of the right arm of LGIII, eDp6, was isolated by J. Hodgkin in association with the deficiency eDf2 (Genetics 96. 649-664, 1980). We have used eDp6 extensively in the mosaic analysis of tra-1 and have made several observations that are consistent with this duplication being a ring chromosome. The strongest evidence comes from the generation and analysis of deletions in the duplication. vab-7(e1564)dpy-18(e364)unc-64(e246);eDp6 hermaphrodites were irradiated with 1500 rads of gamma rays. Of 5500 F1 duptication- bearing progeny, 13 were identified that contained altered duplications (see figure). Altered duplications that failed to complement two of the three markers and duplications that railed to complement a single marker were generated at similar frequencies, suggesting that partial deletions of the duplication were the primary mutational event. Evidence that eDp6 is circular comes from the deletion events of classes D and E in the figure. These classes, which arose at about the same frequency as the others, could have arisen from a single event in a circular chromosome but would require two independent events in a linear chromosome. Additional circumstantial evidence for circularity is that simple recombinants between the duplication and LGIII are not recovered, suggesting that recombination either does not occur or is lethal. However, we have recovered a complex recombinant, suggesting that recombination can occur. The phenotype of the complex recombinant was identical to that of strain CB1517, which is homozygous for eDf2 and carries an undetermined number of copies of eDp6 (Hodgkin, op. cit.). Complementation analysis of the complex recombinant showed that the resulting LGIII is deficient for vab-7, unc-32, suggesting that the recombination resulted in a deficiency of this region, similar to eDf2. That complex recombinants can be recovered and simple recombinants are not suggests that the latter events are lethal. Although simple recombinants between eDp6 and the chromosome have not been recovered, recombinants between ctDp2 (a class A isolate; see figure), and LGIlI are frequent. eDp6 is meiotically unstable; it is recovered in only 50% of the self progeny of duplication hermaphrodites. Mitotically the duplication is stable (loss/cell division is about 1/1000-5000). ctDp2, which covers vab-7, unc-64 is recovered in only 15% of self progeny and is lost mitotically at a rate of about 1/250 cell divisions. The class B isolate, which complements all three markers, is recovered in only 25% of self progeny and is lost mitotically at an estimated 1/100 cell divisions ( not carefully determined). These results suggest that either a segregator function on the duplication has been deleted or that the change in structure or size of these derivatives has a profound effect on segregation (e.g., perhaps ring duplications are mitotically more stable than linear duplications). The circumstances in which eDp6 was isolated (Hodgkin op. cit.) suggest that this duplication and the accompanying deficiency were generated by a single intrachromosomal recombination event. This could have generated a circular free duplication and a deletion chromosome capped by a telomere.