Worm Breeder's Gazette 10(2): 125
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
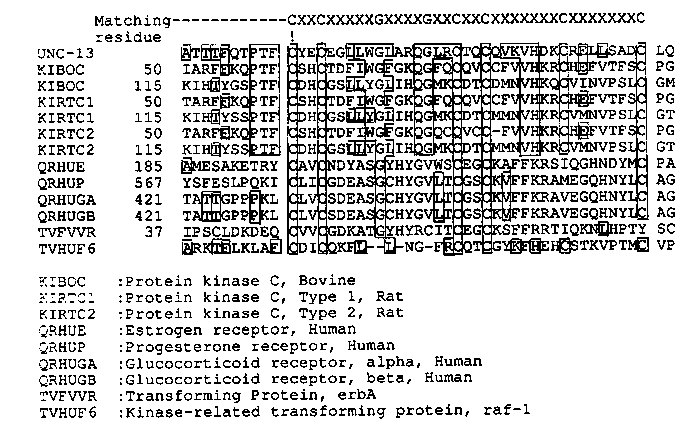
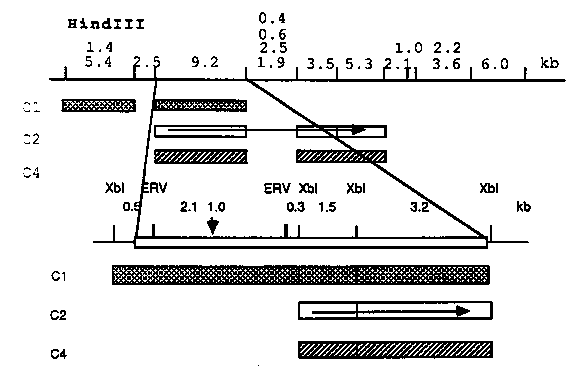
Analysis of unc-13 cDNA clones The identification of unc-13 gene has been described in the last WBG. Since then we have looked for the cDNA clone in 'Ahringer cDNA library 4: N2 mating population', using the polymorphic DNA fragment as a probe, and have found some positives all of which have about 3 kb- long inserts. This is consistent with the result of Northern analysis of the transcript using cosmid clone as a probe although we have not detected the transcript on Northern blot of total RNA using purified polymorphic DNA and cDNA as probes for unknown reason, but probably due to a low copy number of the transcript and/or a large size of the gene. We are now working on Northern analysis of the gene transcript using much more purified mRNA. In order to confirm that the cDNA clones derived from the unc-13 gene, we hybridized cDNA to the genomic Southern blot as well as to the cosmid and lambda genomic clone DNA encompassing unc-13 region. All of these clones hybridized to the polymorphic fragment on the unc- 13 mutant genome and have been localized on the restriction map. At least three different transcripts (more and more coming up) have been identified so far, as shown below in Figure 1. These clones do precisely overlap each other in the region of polymorphic band (shown by a vertical arrow in Figure 1) of unc-13 mutant and encoded in the segment of about 35 kb in length (What a huge gene for only 3 kb mRNAs!). Sequencing one of the cDNAs The whole insert of one of the cDNA clones, C2, was sequenced. The cDNA clone has 2541 bp-long insert which encodes a full 5' region including noncoding stretch of 51 bp in length, but terminates at EcoRI site inside of coding region because there is neither stop codon nor AT-rich 3' noncoding segment. By searching NBRF library we found that amino-terminal half of 'C2 product' is homologous (50%) to protein kinase C (PKC), but not in the carboxy-terminal half of PKC which has a kinase activity. This might mean that 'C2 product' is similar to the regulatory domain of PKC but does not have a kinase activity. It is interesting that the amino-terminal half homologous to PKC contains the typical cysteine-repeating region. The function of the segment in PKC is not known, but the positions of cysteine residues is almost perfectly conserved in DNA-binding domain of steroid hormone receptors and so on (Fig. 2), suggesting that C2 product may be under a similar regulation to PKC in the signal transduction process, and may presumably regulate gene expression of some other genes through DNA-binding. This possibility remains to be elucidated. I am now on the way to finish sequencing a bit of C-terminus of 'C2 transcript' by sequencing of genomic DNA as well as analyzing some other transcripts. I would like to thank Julie Ahringer in Judith Kimble's lab for providing the cDNA library.