Worm Breeder's Gazette 10(2): 123
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
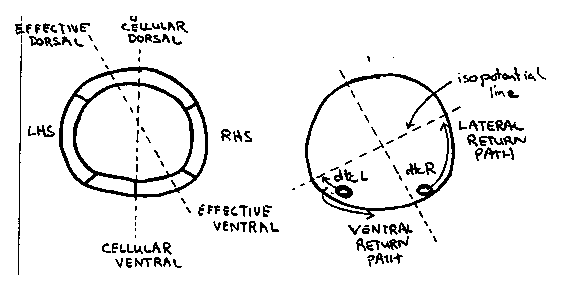
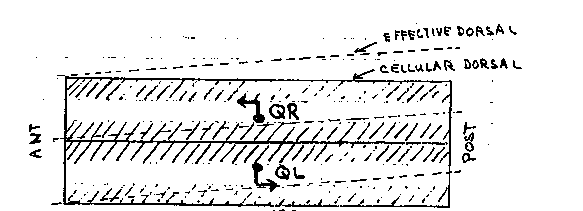
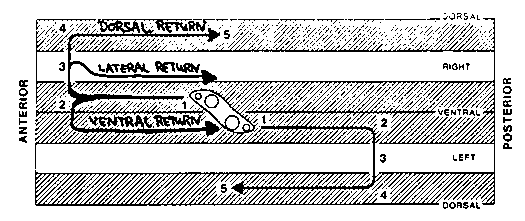
Beneath their smooth exteriors, nematodes have many developmental asymmetries in cell and axon migrations. For example, the Q neuroblasts and the hermaphrodite distal tip cells migrate anteriorward along the right side of the body wall but posteriorward along the left side. The ventral nerve cord is also asymmetrical in that the right fascicle is substantially larger than the left fascicle. This results, in part, from the asymmetric growth of the AVG pioneer axon along the right side of the ventral midline and its recruitment of later axons (R. Durbin and J. Thomson, 1987 CSH Abstracts, p.72). In wild type hermaphrodites, the right and the left distal tip cells first migrate anteriorward and posteriorward, respectively, along the ventral body muscles. Later, they turn dorsalward along the body wall, and finally, return posteriorward and anteriorward, respectively, along the dorsal body muscles (Figure 1). In unc-5 and unc-6 mutants, distal tip cells fail to migrate dorsalward and return along a ventral or lateral path instead. Cells which follow a ventral path are easily recognized because the advancing distal gonad displaces the dark intestine dorsalward. A curious observation is that distal tip cells, more frequently follow a ventral return path when on the left side of the body wall than on the right side (Figure 2). The 'better' performance of right (or anterior) distal tip cells might reflect the presence of a second guidance gradient in the head which is unaffected by these mutations. Alternatively, it could reflect a second gradient present throughout the animal which is tilted (dorsal leftward/ventral rightward) relative to the dorsal/ventral axis defined by hypodermal cell boundaries (Figure 3a). In the absence of the unc-5 and unc-6 products, distal tip cells might respond only to this residual, asymmetrical gradient (Figure 3b). A second asymmetry appears in unc- 40 mutants. In wild-type first stage larvae, the left and right rows of ventral hypodermal cells (Pn) interleave to form a single row In unc-40 mutants, and less frequently in unc 6 mutants, these cell intercalations sometimes fail. The frequency of failed intercalation appears to be about two-fold higher for cells on the right side of the body wall than those on the left side. The non-intercalated Pn cells may later undergo normal cell lineages to form ectopic neurons and lateral vulva-like protrusions which interrupt the row of seam cells. Such protrusions were mistakenly reported to arise from seam cells at the last CSH meeting.) Cells and axons which migrate asymmetrically in wild-type animals may be guided primarily by the conjectured tilted gradient. For example, the AVG axon may follow the ventral mid line rather than the anatomical midline. The assymetric migrations of the Q neuroblasts and distal tip cells could be explained if the tilt increases from anterior to posterior along the length of the animal. ( Figure 4) A single instruction (grow dorsalward) would then guide cells on the left and right sides in opposite directions along the body wall. [See Figures 1-4]