Worm Breeder's Gazette 10(1): 64
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
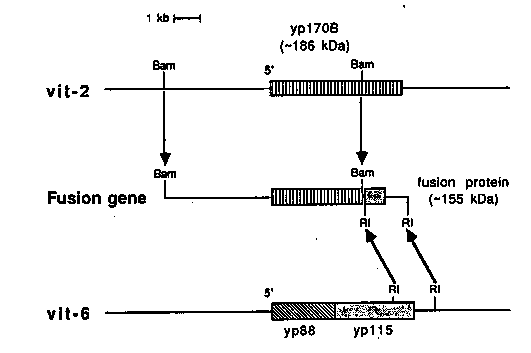
The fusion gene shown below was incorporated onto a plasmid containing sup-7 and injected into tra-3 oocyte nuclei, ala Andy Fire. We obtained three stable transgenic lines containing multiple copies of the plasmid integrated as a tandem array. One strain (temporarily designated 893) has two to four copies of the plasmid DNA and is not viable as a homozygote, while the other two strains (899 and 910) each have about 15 copies per haploid genome. Surprisingly, both of the higher copy strains are viable as homozygotes. The fusion genes appear to be expressed with the same temporal, tissue and sex specificity as the endogenous vitellogenin genes, indicating that sufficient DNA is present to specify correct developmental regulation. The 155 kDa polypeptide product of the fusion gene, which is made in abundance, is not taken up into oocytes, but instead accumulates in the body cavity, and perhaps also in the intestine. It appears to be secreted, but not as well as normal vitellogenins. [See Figure 1] On Western blots the fusion gene product is bound by antibodies specific for the endogenous yolk proteins, yp170 and yp115, but not by those for yp88, demonstrating for the first time that yp115 is at the C terminus of the vit-6 gene product. In each transformed strain, the level of expression from the fusion genes is roughly proportional to the number of copies. That is, homozygous 910 and 899 animals make more 155 kDa fusion protein than heterozygous individuals, which in turn make considerably more than 893 heterozygotes. Interestingly, in both strains containing about 15 copies/haploid genome the level of mRNAs for all of the endogenous vitellogenins drops dramatically (as much as 80% reduction). This has been demonstrated by Northern blots, by in vitro translation and by in vivo synthesis. Furthermore, the level of endogenous vitellogenin synthesis is more severely competed in the homozygotes than in the heterozygotes. The total vitellogenin mRNA accumulation in these transgenic strains remains approximately the same as wild-type except fusion gene mRNA is present in place of the normal mRNA. It appears that 2-4 copies is not enough to compete since strain 893 makes normal levels of vitellogenin mRNAs in addition to the mRNA for the fusion gene. Since there are six endogenous vitellogenin genes, it is remarkable that the presence of only 15 additional genes results in such a severe reduction in production of endogenous vitellogenin mRNA. There are several possible explanations of these results: 1. The extra promoters may be competing for a limiting activator protein. 2. The transcriptional capacity of the intestine may be saturated. 3. The fusion protein accumulating in the intestine may result in destablization of endogenous vitellogenin mRNA.