Worm Breeder's Gazette 10(1): 62
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
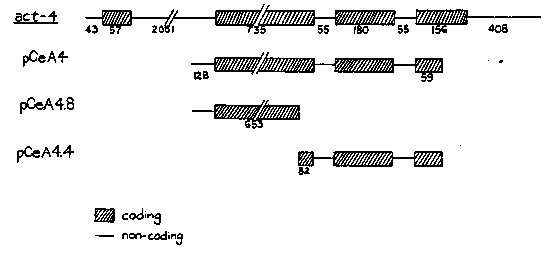
Madl and Herman (Genetics 93: 393-402, 1979) demonstrated that C. elegans is sensitive to slight differences in the balance between the number of X chromosomes and sets of autosomes (X/A ratio). Their experiments showed that 4A;3X animals (X/A=0.75) were fertile hermaphrodites, while 3A;2X worms (X/A=0.67) were normal males. Further, the presence of X duplications in such males resulted in hermaphroditization, the severity of which was proportional to the size of the duplication. These experiments led to the hypothesis that there are numerous sites (at least 4) on the X chromosome which are somehow 'counted' and compared to the number of autosomal sets during sex determination in the developing animal. To determine the nature and distribution of these X-linked sites, we have begun a series of microinjection experiments in which cloned pieces of the X chromosome are assayed for their ability to influence sex determination. Briefly, X-linked clones are injected into the mitotic gonad ( Stinchcomb et al., Banbury Report 20: 251-263, 1985) of 4A;4X hermaphrodites. These animals are mated to diploid (2A;1X) males carrying a small duplication of the X (mnDp8). In the absence of injected X chromosomal material, the duplication is sufficiently small that there is no effect on sexual development; the duplication appears to increase the sensitivity of the animal to the injected DNA, however. In the first experiments, a pool of DNA representing approximately 120 kilobases of the X was injected. Several of the injected worms produced broods which included no males among cross progeny. We inferred from this result that the males may have been pushed over the threshold from male to hermaphrodite development by the injected DNA. One plasmid, pCeA4, obtained from Mike Krause and containing part of the X-linked actin gene act-4 [See Figure 1] in pBR322, was selected for further study. Mosaic intersexes and abnormal males are observed among the 3A;2X progeny of worms injected with this plasmid. Abnormal males are those in which some sexually dimorphic characteristics are poorly developed, e.g. incomplete gonads and poorly developed tails. By contrast, mosaic intersexes possess normal or nearly normal characteristics of both sexes, typically an almost normal male tail and body morphology and a hermaphrodite gonad, often containing embryos and occasionally a vulva. The two classes occur with approximately equal frequencies and represent one-sixth to one-tenth of the 3A;2X cross progeny. In control experiments, injection of either pBR322 alone or a clone of the autosomal act-3 gene led to neither abnormal males or intersexes among over 100 2X;3A progeny. We conclude that the X-linked DNA in pCeA4 contains a sequence (or sequences) which is recognized as being X-linked and therefore influences the assessment of the X/A ratio. Since more than 90% of the bases in the coding regions of act-3 and act-4 are identical and pCeA4 contains neither 5' nor 3' non-coding regions, the most likely candidate for the sequence(s) is within the introns, which are not conserved. We are currently attempting to define further the X-linked sequence involved by injecting the subclones of pCeA4 indicated in the figure. Preliminary results rule out the plasmid containing the two small introns and implicate the regions containing part of the large intron. In addition, we are extending our studies to examine other X-linked genes such as myo-2 to determine the frequency and sequence conservation of other hermaphroditizing sites on the X-chromosome.