Worm Breeder's Gazette 10(1): 21
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
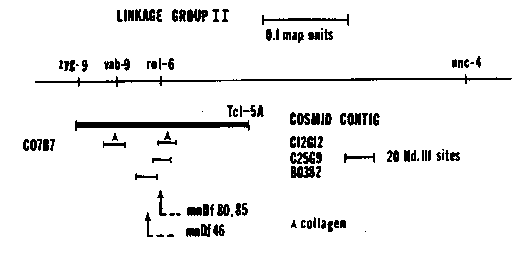
Our lab is interested in the interaction of collagen molecules in the assembly of the cuticle. Since the cuticle is composed mainly of collagen, it is reasonable to assume many of the genes affecting cuticle morphogenesis will be collagen genes. rol-6 is a developmentally regulated genetic locus affecting cuticle morphogenesis. Mutations in rol-6 result in the formation of right handed, helically twisted cuticles. There are dominant, recessive and null alleles of rol-6. The dominant allele has a more severe phenotype. Null alleles are essentially wild type. We have developed several lines of evidence to strongly suggest that we have isolated rol-6 and that it is a collagen gene. A cosmid contig (courtesy of A. Coulson) has been established with the left end anchored by zyg-9 and the right end by a cloned Tc1 element (thanks to J. Park). This contig spans a distance greater than 300 kb and includes rol-6. Southern blots of EcoRI digested cosmid DNAs probed with the cloned collagen gene, col-2, reveal the presence of two collagen genes. Both genes are on 4 kb EcoRI fragments from non-overlapping cosmids (C12G12 and C07B7). Taking advantage of the many deficiencies around rol-6, we have mapped the collagen gene in C12G12 to rol-6 using deficiency strains. Since homozygous deficiency chromosomes are lethals, we have selected mnDf45/mnDf80 for use in mapping rol-6. mnDf45/mnDf80 is a viable strain in which each homologue of linkage group II carries a slightly different, overlapping deletion spanning rol-6. As a result, rol-6 is entirely deleted from these animals. Hybridization of C12G12 to blots of EcoRI digested mnDf45/mnDf80 show that the 4 kb EcoRI fragment containing the collagen gene is absent in this genome. This is the expected result if the collagen gene in C12G12 is rol-6. The collagen gene in C07B7 is present in the mnDf45/mnDf80 genome. The size of the overlap between mnDf45 and mnDf80 is between 7 kb and 17 kb. We are making more precise measurements of the size of the overlap. In addition, C12G12 contains one of the ends for mnDf80 and one for mnDf85. Both of these ends are located in an 8 kb SalI fragment containing the putative rol-6 gene. A cosmid (B0382) to the left of C12G12 contains one of the ends for mnDf46. These ends are probably the left ends of these deficiencies. We have not been able to identify either end of mnDf45. Cloning of the dominant allele, su1006, and the recessive allele, e187, of rol-6 is presently being done. Sequencing of the wild type allele is under way. We have preliminary results from Northern blot analysis using a 2.2 kb HindIII fragment containing the putative rol-6 gene as a probe to egg, L2 to dauer, L3 to L4 and L4 to adult molt total RNAs. A 1.2 kb transcript is seen at the L2 to dauer molt. A 1.2 kb and a 1.0 kb transcript are detected at the L3 to L4 and L4 to adult molts. No transcript is observed in total egg RNA. These blots were washed under high stringency conditions (.03 M Na+, 10% formamide, 65 C). The time of expression of the putative rol-6 gene agrees with the time of appearance of the rol-6 phenotype. We cannot rule out the possibility of cross hybridization between rol-6 and other collagen gene transcripts. Total L4 to adult RNA from the mnDf45/mnDf80 strain has been prepared and Northern blots will be done. If the transcripts are specific for rol-6 then we expect that the 2.2 kb HindIII probe will not hybridize to mnDf45/mnDf80 total RNA. Genomic DNAs have been made from several putative null alleles of rol-6 (provided by R. Horovitz). Southern blots of HindIII, EcoRI and BamHI triple digested genomic DNAs from these strains have been made and probed with either the 2.2 kb HindIII or an 8 kb SalI fragment containing the putative rol-6 gene and flanking sequences. Triple digests were done to facilitate detection of subtle differences In the length of restriction fragments associated with rol-6. Initial results have not shown any detectable differences between these null strains and N2. There are still several null genomic DNAs to be examined. Attempts at Tc1 tagging rol-6(e187) by crossing e187 into the HH*6 mutator background and screening for non-rollers have not been successful (HH*6 supplied by M. Finney). However, a semidominant, unlinked suppressor of rol-6(e187) has been isolated from this screen and backcrossed six times into N2. Suppression of rol-6 e187) is complete in the homozygous state (e187/e187; sup/sup). As a heterozygote (e187/e187; sup/+), worms roll as L3s but not as adults. We have not observed any revertants while maintaining the suppressor in the HH*6 background. Genomic DNAs have been prepared from the backcrossed suppressor strain and Tc1 screening is underway. The suppressor is being mapped genetically. The collagen gene in C07B7 may be vab-9, y close to the left of rol-6 and the vab-9 phenotype could be a cuticle defect. We are presently subcloning this collagen gene and beginning to do expression studies and molecular mapping. [See Figure 1]