Worm Breeder's Gazette 10(1): 126
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
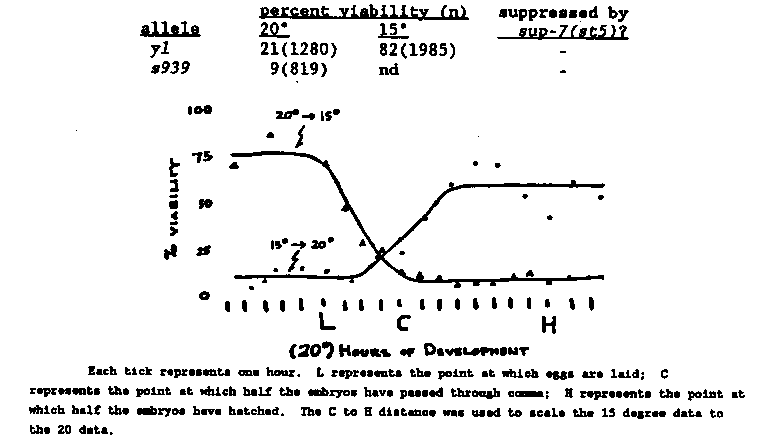
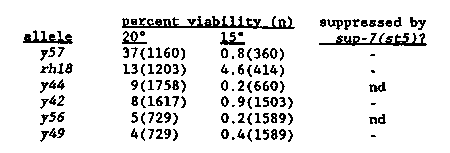
Mutations in dpy-27 and dpy-28 affect the viability of XX but not XO animals. In addition, these mutations disrupt dosage compensation resulting in XX but not XO animals over-expressing their X-linked genes (as assayed by Northern analysis [Meyer and Casson, Cell 47:871 1986] and by morphogenetic assay [DeLong, et al. Genetics, in press]). We have recently isolated five new dpy-27 alleles as suppressors of xol-1 XO-specific lethality (Miller, et al. this WBG). All are very similar in phenotype. XO animals appear unaffected, homozygous XX progeny of heterozygous mothers are dumpy but viable, and most homozygous XX progeny of homozygous mothers are inviable. Both the dumpiness and lethality of dpy-27 mutations are zygotically rescuable. None of the alleles yet tested are suppressed by the tRNA amber suppressor sup-7(st5) for either dumpiness or lethality. [See Figure 1] dpy-27 mutant animals which reach adulthood do so between four and seven days after fertilization. The majority of the inviable animals hatch and persist as L1 larvae for several days. Examination by Nomarski optics reveals no obvious morphological defects (besides a dumpy phenotype) in these animals; however, their guts tend to be devoid of bacteria and they appear to be starving. In addition it is intriguing that all six alleles are cold-sensitive for this lethal phenotype; mutations in dpy-27 may reveal a cold sensitive process. We are using y57 to define the temperature sensitive period of this lethality. Mutations in dpy-26 (which have a similar phenotype to dpy- 27 mutations) are also cold-sensitive for their associated maternal- effect XX-specific lethality (L. DeLong, personal communication). By reverting dpy-27(y57) at the restrictive temperature it may be possible to define genes involved in this putative cold sensitive process. The two alleles of dpy-28 also have a similar phenotype to those described above for dpy-27, with the exception that homozygous XX progeny of heterozygous mothers are not dumpy, but are rather egg- laying defective (some animals are slightly dumpy). As with dpy-27, most homozygous dpy-28(y1 or s939) XX progeny of homozygous mothers are inviable, the survivors being dumpy. Again, both the dumpiness and lethality of dpy-28 mutations are zygotically rescuable. These zygotically rescued heterozygotes are essentially wild-type, unlike dpy-28 homozygous progeny of heterozygous mothers. This suggests that the zygotically provided dpy-28 product is superior (either quantitatively, qualitatively, or temporally) to that provided maternally. In addition, both y1 and s939 are low level him mutations. [See Figure 2] Preliminary experiments suggest that s939 is unaffected by temperature. y1 and s939 are not suppressed by sup-7(st5) for either dumpiness or lethality. The temperature sensitivity of y1 has allowed us to define a brief temperature sensitive period for the lethality which is approximately coincident with the comma stage of embryogenesis. [See Figure 2] The upshift curve has an inflection point during embryogenesis, suggesting that the maternal endowment of y1 product is temperature labile. If the maternally provided y1 product were temperature stable one might expect that upshifting embryos would have no effect on viability; the resultant curve would be flat within the time period of embryogenesis. Alternatively, an upshift curve such as we see could result from the maternal endowment of 15 C y1 product being insufficient to rescue embryos. As already discussed, there is reason to believe that zygotically provided dpy-28 product is superior to maternally provided product. We are presently performing temperature pulse experiments to determine if comma stage is indeed the specific critical period. What is the relationship between this lethality and the known over- expression of X-linked genes in dpy-28(y1)? At the moment we do not know, but several possibilities exist. The lethality may be a direct or indirect consequence of the role dpy-28 plays in dosage compensation, or the lethality may be due to a requirement for dpy-28 independent of this role. But whichever is the case, it is clear that dpy-28 is required during this period of embryogenesis, and that lack thereof results in lethality.