Worm Breeder's Gazette 10(1): 119
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
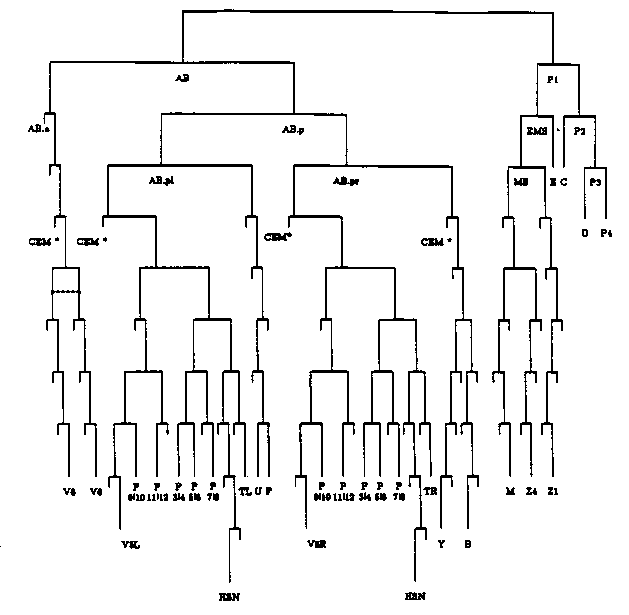
We are analyzing genetic mosaics of the sex determination gene tra-1. We would like to know if the sexual fates of cells in C. elegans are autonomously determined or if they are influenced by interactions with other cells. The strain from which genetic mosaics are isolated is homozygous for tra-1 (either e1099 or e1781) and carries the free duplication eDp6 which provides tra-1 + activity. The self progeny from this strain are primarily duplication-bearing hermaphrodites and non-duplication-bearing XX Tra males (there are subtle differences in the Tra phenotypes of e1099 and e1781 males). Rare somatic loss of the free duplication should result in an intersexual animal that expresses some hermaphrodite and some male fates. If tra-1 determines sexual phenotype cell-autonomously, then somatic loss of the free duplication will result in intersexual animals that express male fates only in cells that are lineally related. If some sexual phenotypes are determined non-cell-autonomously, then somatic loss will result in intersexual animals that express male fates in cells that may not be lineally related. 60,000 progeny from tra-1(e1099 and e1781) eDp6 hermaphrodites were scored for sexual phenotype in a screen for genetic mosaics that had lost the duplication in an early cell division. Among these about half appeared to be hermaphrodites, indicating retention of eDp6. Two intersexual animals were identified in this screen whose phenotypes can best be interpreted as caused by loss of the free duplication in an early cell division. One intersex appeared to have lost the duplication in the AB cell and retained it in the P1 cell. This animal had a complete male tail [fan, 18 rays, crumpled spicules (male sex muscles are required for proper shaping of the spicules)], a hermaphrodite gonad, hermaphrodite sex muscles, and a vulva (herniated). The animal was self-fertile and produced six progeny, three males and three hermaphrodites, demonstrating that the duplication was present in the germline. The tra-1 allele was e1099. The germ line, the gonad, the sex muscles, and the intestine are all derived from the P1 cell (see figure) and in this animal were hermaphrodite. The tail structures listed are all derived from the AB cell and in this intersex were male. This phenotypic pattern, except for the induction of the vulval cells, is consistent with cell- autonomous function of tra-1.A second similar animal was isolated from the e1781 stock. However, we could not identify the first ray on either side (ray cell groups were not scored) , and at least some of the male-specific sex muscles were present. Consistent with the presence of male sex muscles, the spicules were not crumpled. This animal was also self-fertile and produced three male progeny, an insufficient number of progeny to rule out presence of the duplication in the germ line. This phenotypic pattern could be the result of first-cleavage duplication loss in AB and a second later loss in the derived lineage leading to or including the sex myoblast M. In a preliminary screen (13,000 duplication-bearing progeny) for later duplication loss events we identified an intersex that we interpret as a first-cleavage duplication loss in P1. This animal had a male gonad and male sex muscles, with a hermaphrodite body and tail. There was no vulva. The gonad contained one oocyte and many mature sperm. (e1781 Tra males often produce oocytes.) The intestine did not produce yolk protein (assayed by whole-mount staining with monoclonal Ab OIC1). In this animal, the opposite of the first intersex discussed, all structures derived from the P1 cell were male ( germ line, gonad, sex muscles, and intestine). The male tail structures listed above for the first intersex and derived from the AB cell were not present (i.e., were hermaphrodite). This phenotypical pattern is also consistent with cell-autonomous function of tra-1.We have also examined 33 other intersexes that appear to result from duplication loss in later divisions. All are consistent with cell autonomous function of tra-1; that is, each of the observed phenotypic patterns can be explained by a single loss of the duplication in a lineage derived from the AB cell. The sexual phenotypes of these intersexes are listed below; for each is indicated the number observed, the precursor cell(s) that must have been male (not carrying the duplication), and the point(s) of duplication loss that most simply explain(s) the phenotype (see accompanying figure). Three intersexes expressed male hooks. One of these also expressed at least three rays (on the left side of the animal) and had a herniated vulva. This phenotype can result from loss of the duplication between AB.pl and AB.plap, which can generate three T-rays, a single ray from V5 (not seen), and a male P10 cell. Consistent with the first two intersexes described, tra-1 (male) P cells in the presence of a hermaphrodite anchor cell results in a herniated vulva, suggesting that the 'male' cells respond, but not normally, to the inducing signal from the gonad. This is also a frequent phenotype of tra-1 (e1488) animals, which also have a hermaphrodite gonad (anchor cell) in the presence of tra-1 P cells. The other two intersexes that expressed the hook did not generate either rays or a herniated vulva They can be explained by duplication loss in the generation of the P10 cell or in its daughter P10.p. Because the ray generated from V5 may not have been detected, the loss could have occurred earlier. Two animals expressed 8 to 10 rays and no other male traits, suggesting that both V6 cells were male. This phenotype can be caused by the loss of the duplication at AB.appp or in one of the preceding three cell divisions. Seven animals expressed three to four male rays suggesting that a T cell, and occasionally V5 was male. This phenotype can result from duplication loss at one of three divisions in either the AB.pla or the AB.pra lineage. Two animals expressed a male B-cell marker (refractile cuticle associated with the spicules and the gubernaculum) and at least three rays, suggesting a duplication loss at AB.pr. Finally, twenty-one animals expressed the B-cell marker but no other male fates, consistent with the duplication loss in any of the seven cell divisions from AB.pr leading to the B-cell. Based on our results so far (except for the induction of P.np cells by the anchor cell), the sexual phenotypes of tra-1 mosaic intersexes are consistent with cell autonomous determination of sexual identity by the tra-1 gene in somatic tissues. [See Figure 1] [See Figure 2]
